慢病毒(lentivirus)是逆转录病毒的一种,是RNA病毒,需要相对较长的孵育时间,所以称之为“慢”病毒。慢病毒可以感染分裂细胞、感染非分裂细胞,在感染能力方面,可以有效地感染神经元细胞、肝细胞、心肌细胞、肿瘤细胞、内皮细胞、干细胞等多种类型细胞,此外由于其可以整合至细胞基因组,因此也常用于稳定细胞株的筛选。
维真生物拥有十余年病毒包装经验,依托于成熟稳定的慢病毒包装平台和严格的生产质控,可以提供从慢病毒质粒设计、构建、病毒包装、细胞系建立至基因表达分析在内的所有服务外包项目。全套的慢病毒载体和表达系统可用于体内外表达多物种cDNA、lncRNA、circRNA、shRNA和CRISPR/gRNA。

下面是服务内容(具体规格、费用、货期请详询客服):
| 服务编号 | 服务类型 | 规格 | 目录价 | 货期 |
|---|---|---|---|---|
| WZ060001 | 小包装、OE & CRISPR/Cas9 | 滴度≥1×10E8 TU/ml;体积1ml | 询价 | 详询 |
| WZ060002 | 大包装、OE & CRISPR/Cas9 | 滴度≥1×10E9 TU/ml;体积1ml | 询价 | 详询 |
| WZ060003 | 小包装、单shRNA | 滴度≥1×10E8 TU/ml;体积1ml | 询价 | 详询 |
| WZ060004 | 大包装、单shRNA | 滴度≥1×10E9 TU/ml;体积1ml | 询价 | 详询 |
| WZ060005 | 小包装、shRNA(3保1) | 滴度≥1×10E8 TU/ml;体积1ml | 询价 | 详询 |
| WZ060006 | 大包装、shRNA(3保1) | 滴度≥1×10E9 TU/ml;体积1ml | 询价 | 详询 |
*如您对上述服务感兴趣,请您点击“欢迎垂询”留下您的信息,我们将为您提供具体实验方案和项目价格。
|
↓↓ 分以下几个部分做详细介绍 ↓↓ |
慢病毒是一种有包膜的RNA病毒,直径为80-120nm,呈二十面体对称结构、球形。病毒颗粒最外层是包膜(包膜蛋白决定了感染细胞的类型),再往里依次为基质蛋白和衣壳,最里面是两条相同的正股RNA链和酶(逆转录酶、整合酶和蛋白酶)。
下图是慢病毒的结构示意图:

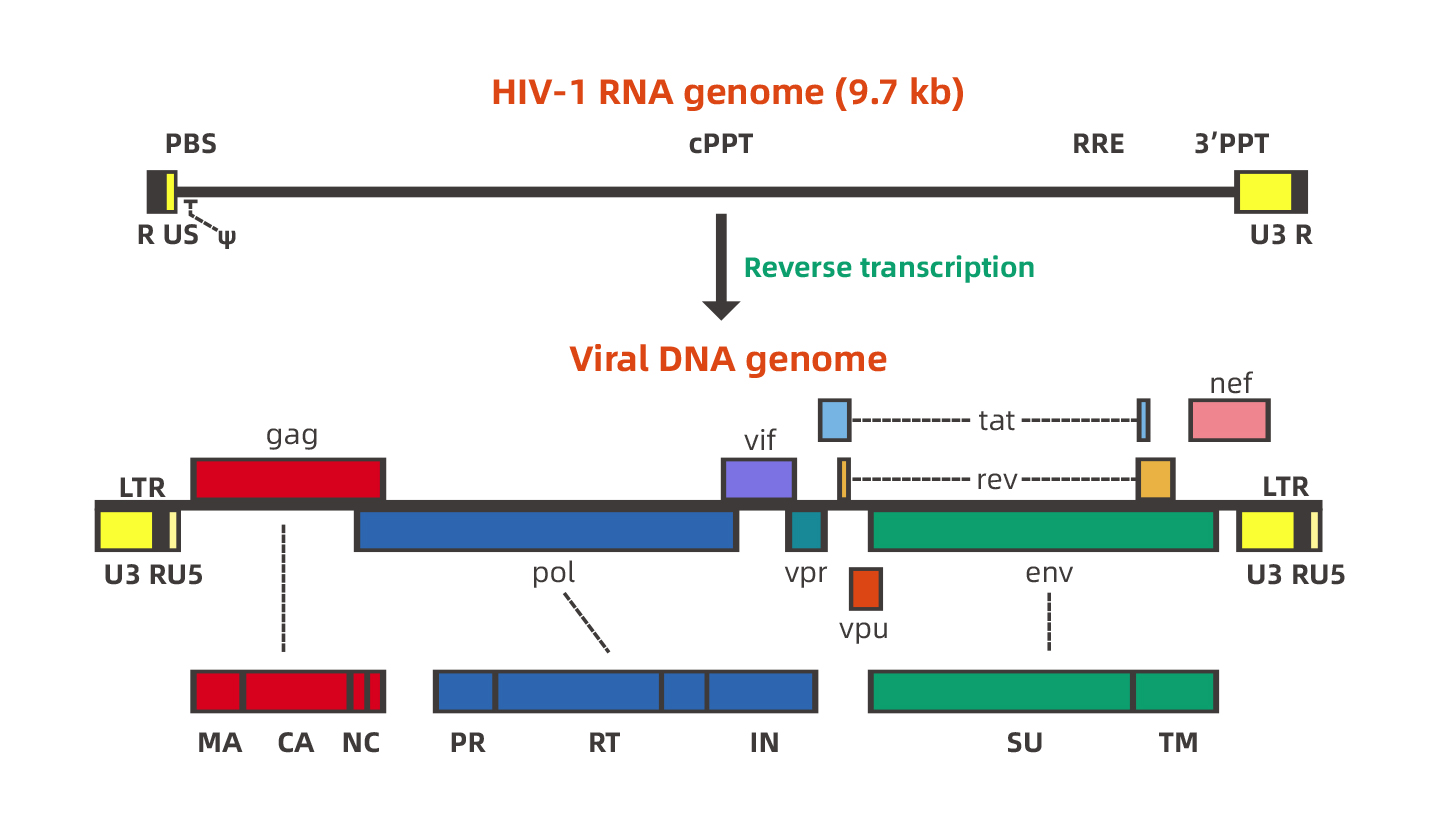
慢病毒含有复杂的基因组。以HIV-1(人类免疫缺陷I型病毒)为例,HIV是一种单链RNA病毒,其基因组长度约为9.7kb。基因组两端各有一个反向末端重复序列(inverted terminal repeat, ITR),中间包括gag、 pol、env 3个结构基因及tat、rev、nef、vif、vpr、vpu 6 个调节基因。下图是HIV-1基因组的示意图:
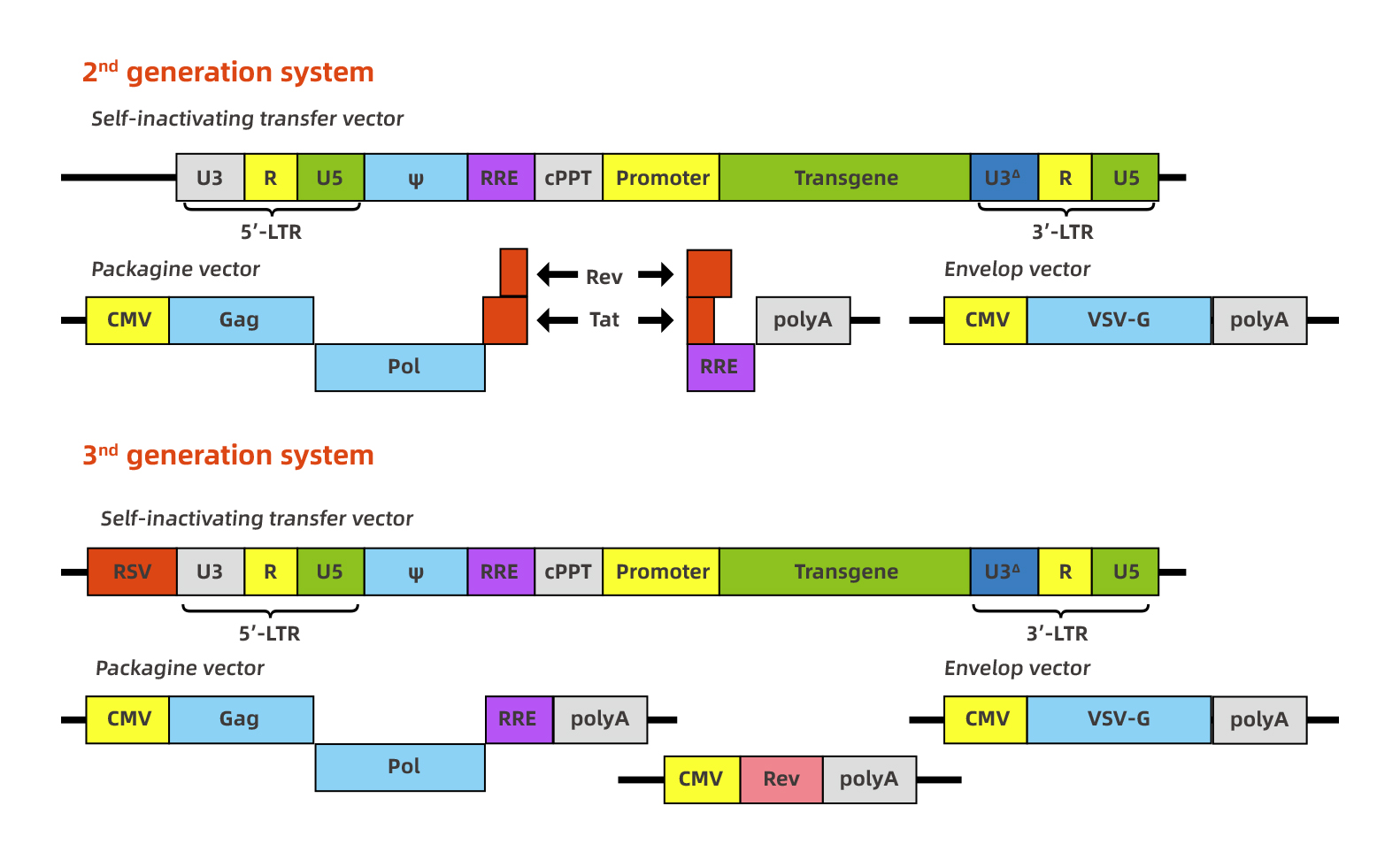
慢病毒是以HIV-1为基础发展起来的基因治疗载体。慢病毒载体的研究发展得很快,研究的也非常深入。出于包装能力和安全性考虑,目前慢病毒载体已发展了很多代,其中2代和3代是目前大家应用比较广的。慢病毒载体的包装容量为6kb,但是通常超过3kb的话会明显影响慢病毒的滴度。下面是2代和3代慢病毒系统的示意图:
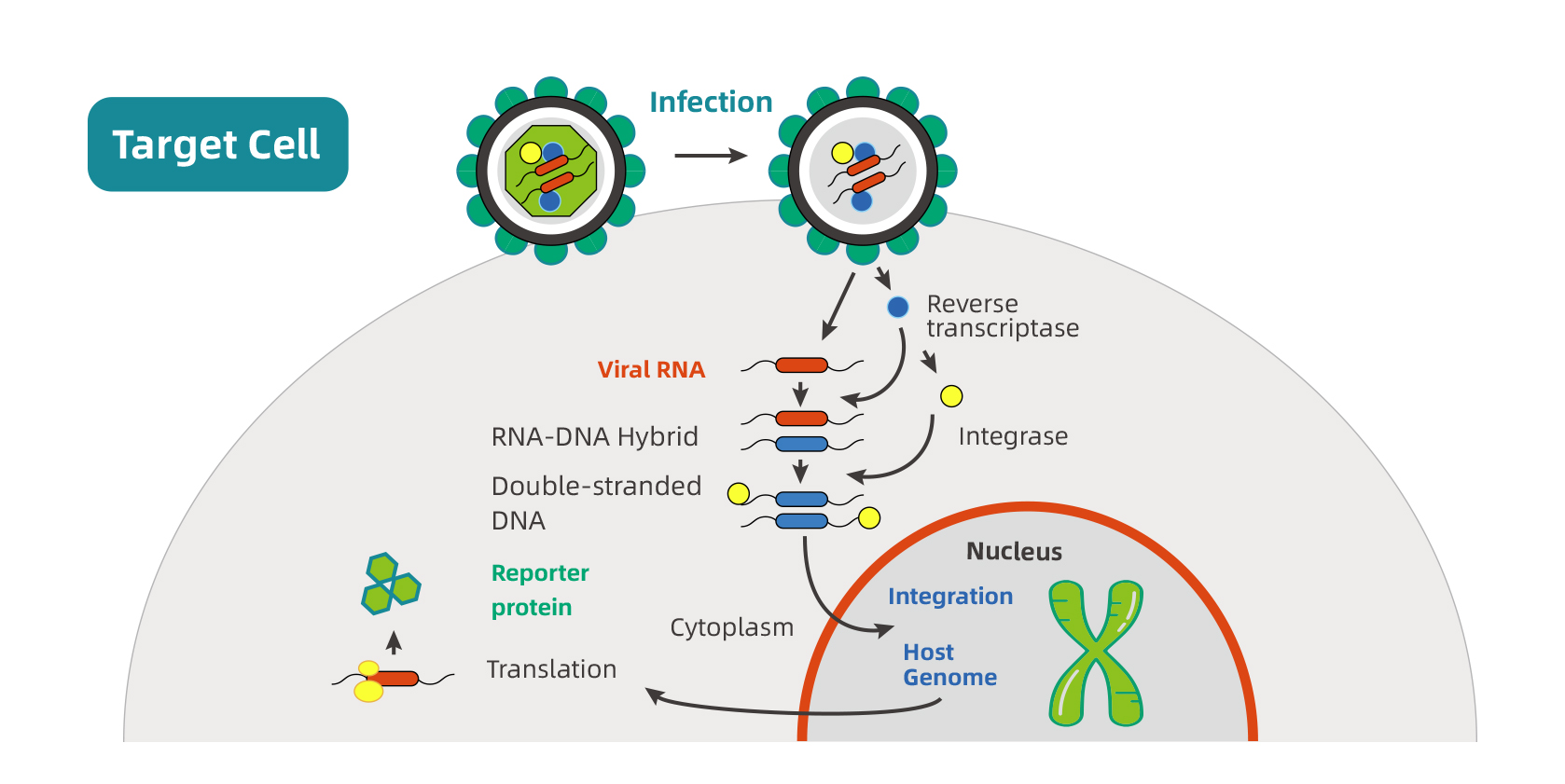
慢病毒感染细胞的第一步是通过包膜蛋白与细胞表面受体结合。慢病毒与宿主细胞膜融合后释放结构蛋白、酶蛋白和病毒核心。病毒RNA在逆转录酶的作用下逆转录并与整合酶形成整合前复合物。整合前复合物进入细胞核后,整合酶催化其整合至宿主基因组。外源基因前启动子驱动其在细胞质中表达。
下图是慢病毒感染细胞的示意图:
1. 慢病毒有广泛的宿主范围,能够有效地感染分裂细胞、非分裂细胞,特别适合于一些难转染的细胞(如原代细胞、干细胞、不分化的细胞)和对腺病毒感染具有较强免疫反应的细胞(如树突状细胞、单核细胞和间充质干细胞)等,能大大提高目的基因转导效率,而且目的基因整合到宿主细胞基因组的几率大大增加。
2. 慢病毒能将外源基因有效地整合入宿主染色体上,介导目的基因稳定、长期的表达。因此,可以利用慢病毒建立稳定细胞株。
3. 适用范围更广,不仅能用于体外实验,还能用于活体动物模型,尤其是动物成瘤实验。
慢病毒应用策略及客户案例分享 |
|||||||||||||||||||
1、慢病毒体外应用策略 |
|||||||||||||||||||
|
▲ 产品:维真shRNA慢病毒 抗性:嘌呤霉素 筛选周期:三周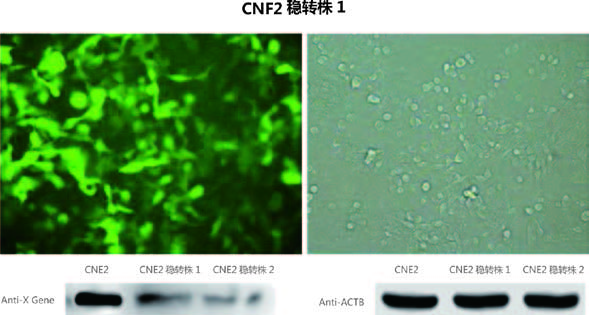
▲ shRNA 载体:pLent-U6-GFP 抗性:嘌呤霉素(Puromycin) 筛选周期:三周 |
||||||||||||||||||
1.1 慢病毒 - 基因过表达 - 乳腺癌作者利用慢病毒在乳腺癌细胞MCF7 和ZR75-1 中过表达piR-36712,发现细胞的增殖、迁移及侵袭都受到 了抑制;此外,体内异种移植实验同样证明了piR-36712 具有抑制乳腺癌细胞恶性表型的作用。
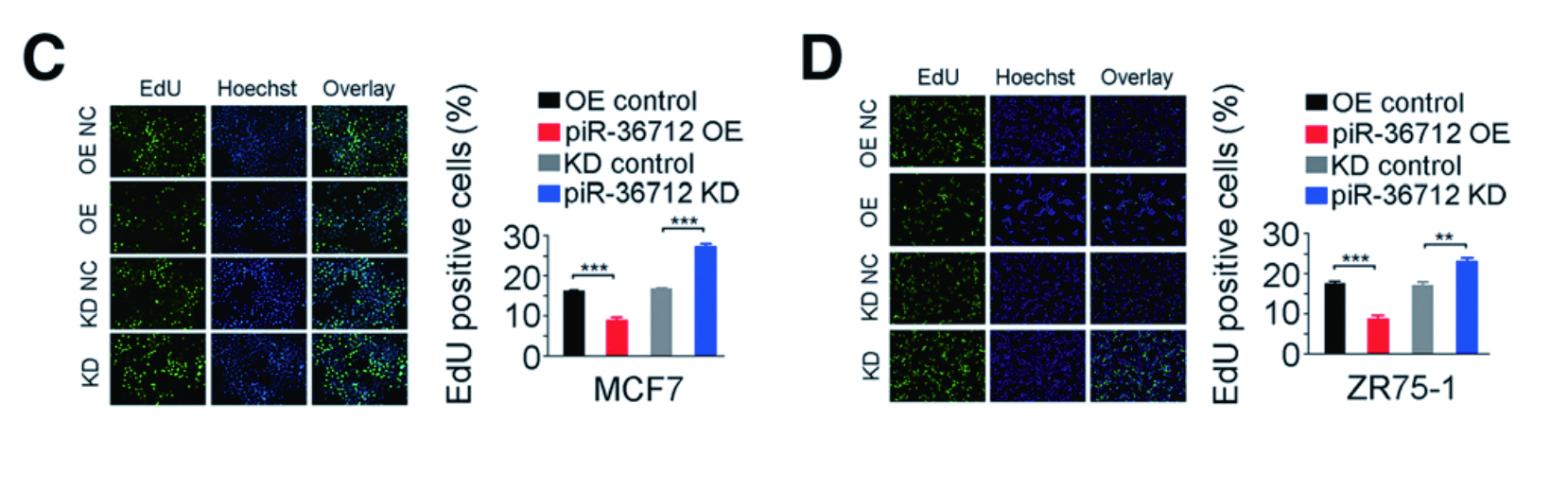
▲ 过表达piR-36712 抑制乳腺癌细胞的恶性表型 |
1.2 慢病毒 - 基因过表达 - 神经 脊髓损伤为探讨CBD-NT3 和TrkC 对体外功能神经网络组织构建的影响,作者设计了未经CBD-NT3 修饰的LOCS 和 未过表达TrkC 的iPSC-NSCs 作为对照组。使用Lv-TrkC(维真生物提供)转染iPSCs,然后诱导其分化为 NSCs,将CBD-NT3 与LOCS 孵育后,发现CBD-NT3 能均匀地结合在LOCS 上。作者在CBD-NT3 修饰的 LOCS 上接种iPS-NSCs- TrkC 构建功能神经网络组织,发现CBD-NT3 修饰的LOCS 可促进iPS-NSCs-TrkC 的存活和增殖。
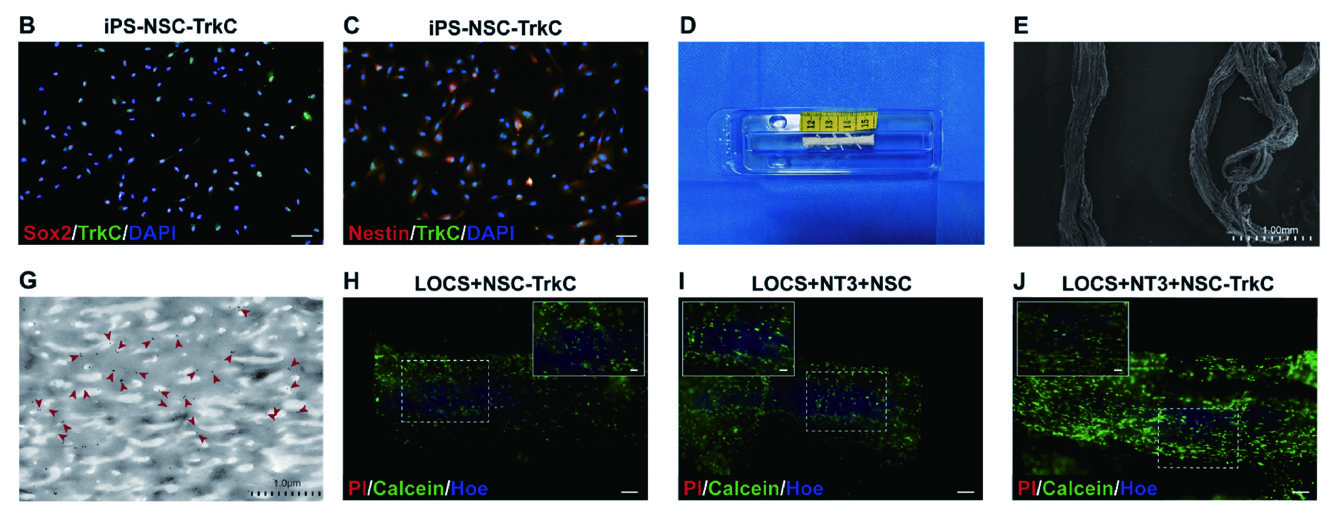
▲ 基于LOCS 和iPSCs 的功能神经网络组织构建 |
||||||||||||||||||
1.3 慢病毒 - 基因沉默 - 纤维肉瘤核酸适体S11e 特异性识别纤维肉瘤并抑制肿瘤生长,作者使用慢病毒敲低S11潜在结合靶点Diablo/ SMAC 的表达,发现S11e 对HT1080 细胞迁移和侵袭的抑制作用被消除,揭示Diablo/ SMAC 蛋白是纤维 肉瘤细胞中S11e 的功能性结合靶点,通过与S11e 相互作用抑制HT1080 细胞的迁移和侵袭。
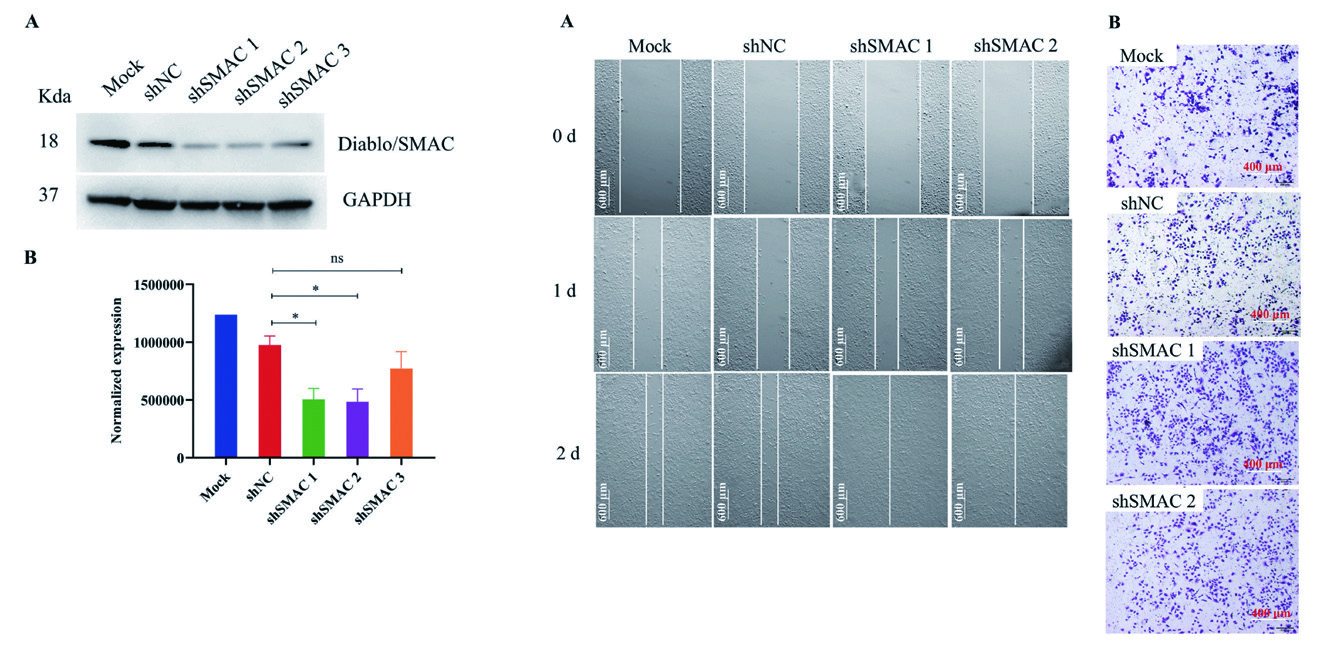
▲ Diablo/SMAC shRNA敲低效果(左)▲ SMAC的表达降低后,S11e对HT1080细胞迁移和侵袭的抑制作用被消除(右) |
1.4 慢病毒 - 基因沉默 - 糖尿病肾病ABCA1 缺失加重肾小球内皮细胞(GEC)炎症损伤和凋亡,促进糖尿病肾病(DKD)的发展。作者利用慢病毒递送shRNA-ABCA1进行体外实验,进一步探讨ABCA1缺失加剧GEC炎症损伤和凋亡的机制。结果表明,ABCA1 敲低能显著上调ERS相关蛋白的表达,并促进凋亡水平的升高。
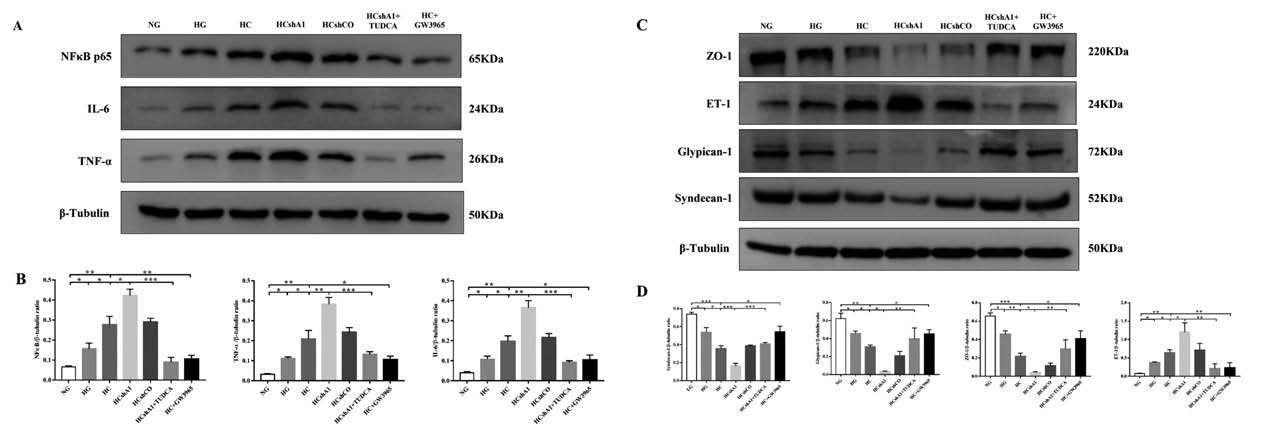
▲ ABCA1 缺失加重了GEC 凋亡 |
||||||||||||||||||
1.5 慢病毒 - 基因沉默 - 胃癌作者将携带AFAP1L1 的干扰慢病毒转染AFAP1L1 高表达的胃癌AGS 细胞系,并在AFAP1L1 低表达的 MKN74 细胞系中利用慢病毒实现AFAP1L1 的过表达。结果发现,AFAP1L1 在AGS 中下调可抑制其增殖, 并且显著降低了AGS 细胞的迁移和侵袭能力;而AFAP1L1 在MKN74 中过表达可显著促进其增殖能力,增加迁移和侵袭能力。此外,体内转移实验表明,AFAP1L1 在AGS 细胞中表达的下调显著抑制了肺和肝转移 灶的形成。
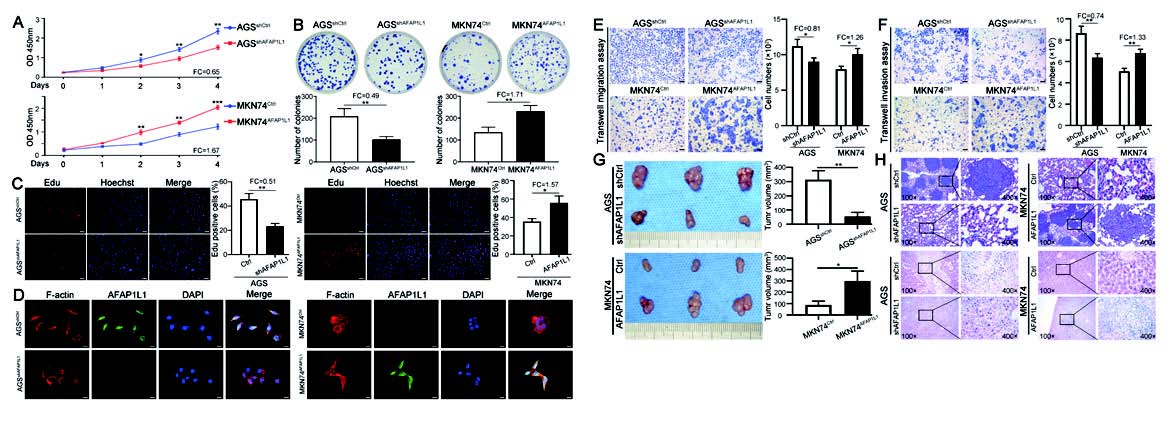
▲ AFAP1L1 的高表达促进GC 细胞体外增殖、侵袭和体内生长、转移 |
1.6 慢病毒 - 基因敲除 - 视网膜血管生成视网膜感光细胞是永久分化的细胞,很难被LV 靶向,当双非整合型慢病毒(NILV) 作为基因编辑载体用于治 疗病理性眼内血管生成( 如PDR) 时,视网膜神经细胞可能受到保护。因此作者开发了一种双NILV 系统, 用于在眼睛玻璃体内递送PE6x,以在小鼠VEGFR2 位点(NC_00071.6) 产生T17967A 突变,从而更早的 停止转录,并且突变的VEGFR2 会阻断VEGF/VEGFR2 信号通路,从而抑制病理性视网膜血管生成。在双 NILV 系统中,LV5 被创建用于表达SpCas9n (H840A) 的融合蛋白,LV6 表达显性失活MLH1 (MLHdn) 和靶 向非编辑链的sgRNA(ngRNA)。
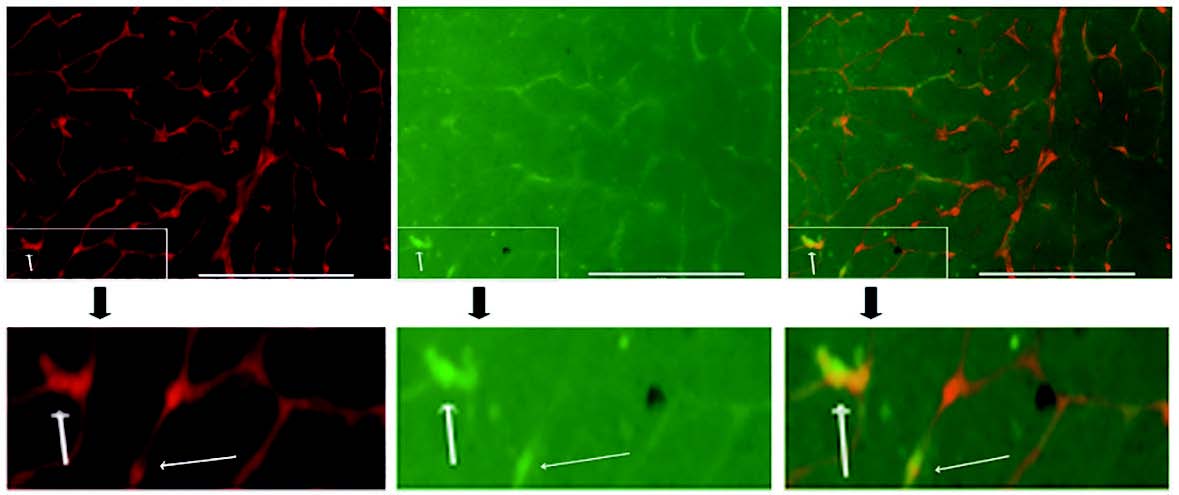
▲ NILVs 在体内转导血管内皮细胞 |
||||||||||||||||||
2、慢病毒体内应用策略(部分) |
|||||||||||||||||||
2.1 慢病毒 - 体内示踪 - 三阴性乳腺癌在肿瘤研究中,通常需要在体内研究肿瘤的发生、发展及转移,此时就需要对其过程进行示踪,研究者可以借助携带荧光素酶(luciferase)基因或荧光蛋白基因的慢病毒感染肿瘤细胞系,构建稳定表达荧光素酶/荧光蛋白的细胞株,并借助细胞诱导肿瘤模型,完成肿瘤发生、发展以及转移等过程示踪。 作者利用携带Luciferase的慢病毒感染MDA-MB-231细胞,建立TNBC异种移植模型,随后进行尾静脉注射FeS2−PEG8纳米晶体。在局部近红外光照射下,发现肿瘤温度显著升高,并在数秒内快速反应,这是 由于注射后FeS2-PEG8 在瘤内积聚造成的。TNBC肿瘤在大小和重量上显著减小,PTT干预的异种移植小鼠的生存期延长,体重稳步增加。
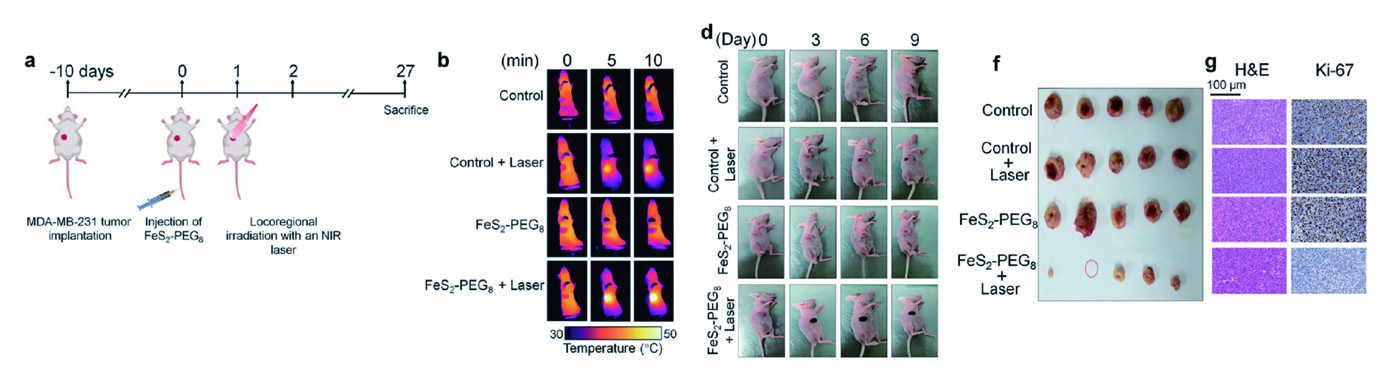
▲ FeS2-PEG8 NCs 抑制小鼠TNBC 发展和骨转移 |
2.2 慢病毒 - 动物成瘤 - 胶质母细胞瘤为探索LILRB2 在胶质母细胞瘤(GBM)中的功能作用,作者利用慢病毒构建了过表达pirb(小鼠体内 LILRB2 的唯一同源基因)的GL261 细胞,然后建立了C57BL/C 小鼠皮下肿瘤模型。与对照组相比,注射 GL261-nc-sEVs 可显著促进GBM 的进展,GL261-pirb-sEVs 的促肿瘤能力明显强于GL261-sEVs。肿瘤样 本中,相比GL261-sEVs 组,在GL261-pirb-sEVs 治疗的肿瘤部位观察到肿瘤浸润的CD8+T 淋巴细胞更少, MDSCs 更多。免疫细胞检测发现,GL261-sEV 和GL261-pirb-sEVs 组CD8 + T 细胞百分率下降,MDSCs 比例上升。以上表明sEV 上的pirb 可以通过诱导MDSCs 发挥作用。
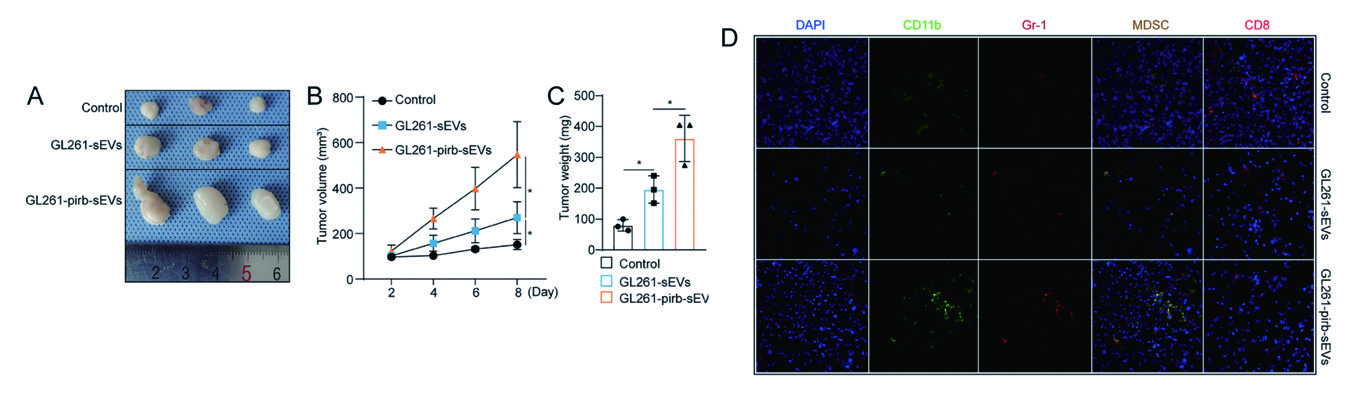
▲ sEVs-pirb 促进GBM 的进展 |
||||||||||||||||||
2.3 慢病毒 - 动物成瘤 - 黑色素瘤作者用干扰慢病毒诱导了ADAR2 敲除的黑色素瘤干细胞中内源性DOCK2 的特异性敲低,并将DOCK2 mRNA 编辑的细胞和DOCK2 mRNA 未编辑的细胞分别注射到裸鼠体内,发现DOCK2 mRNA 编辑明显促进 了小鼠的肿瘤生长,实体瘤的大小及重量显著增加,同时黑色素瘤干性基因表达上调,表明DOCK2 mRNA 编辑可以促进黑色素瘤干细胞在体内的成瘤。
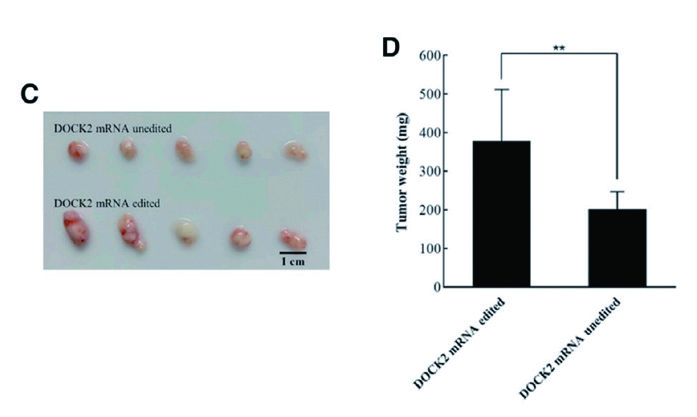
▲ DOCK2 mRNA 编辑可以促进黑色素瘤干细胞在体内的成瘤 |
2.4 慢病毒 - 动物成瘤 - 肾癌为评估SETD8 基因表达下调对肾脏肿瘤生长的影响,研究人员利用慢病毒干扰载体及对照载体建立了 SETD8 表达下调的肾透明细胞癌(ccRCC) 786-O 细胞系及对照。将SETD8-shRNA 或control-shRNA 细胞 植入BALB/c 裸鼠皮下,建立肿瘤异种移植模型。实验表明SETD8 的表达下调明显延长了体内肿瘤生长的 起始时间并降低了其生长速度,文章表明沉默SETD8 基因表达可显著抑制ccRCC 移植瘤的体内生长。
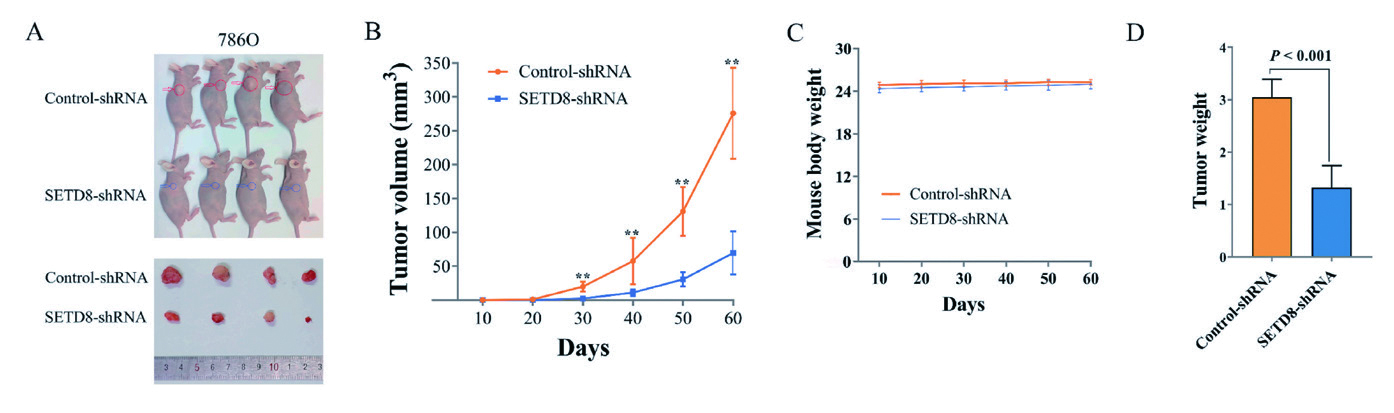
▲ 沉默SETD8 降低ccRCC的致瘤潜能,缩短其在体内的生存概率 |
||||||||||||||||||
|
↑↑ 点击返回,查看其它部分 ↑↑ |
维真生物采用的包装系统是2代包装系统,即三质粒系统,该系统包括1个包装质粒(psPAX2)、1 个包膜质粒(pMD2G)、1个目的基因质粒和1株包装细胞(293T cell)。下图是2代慢病毒包装系统的示意图:
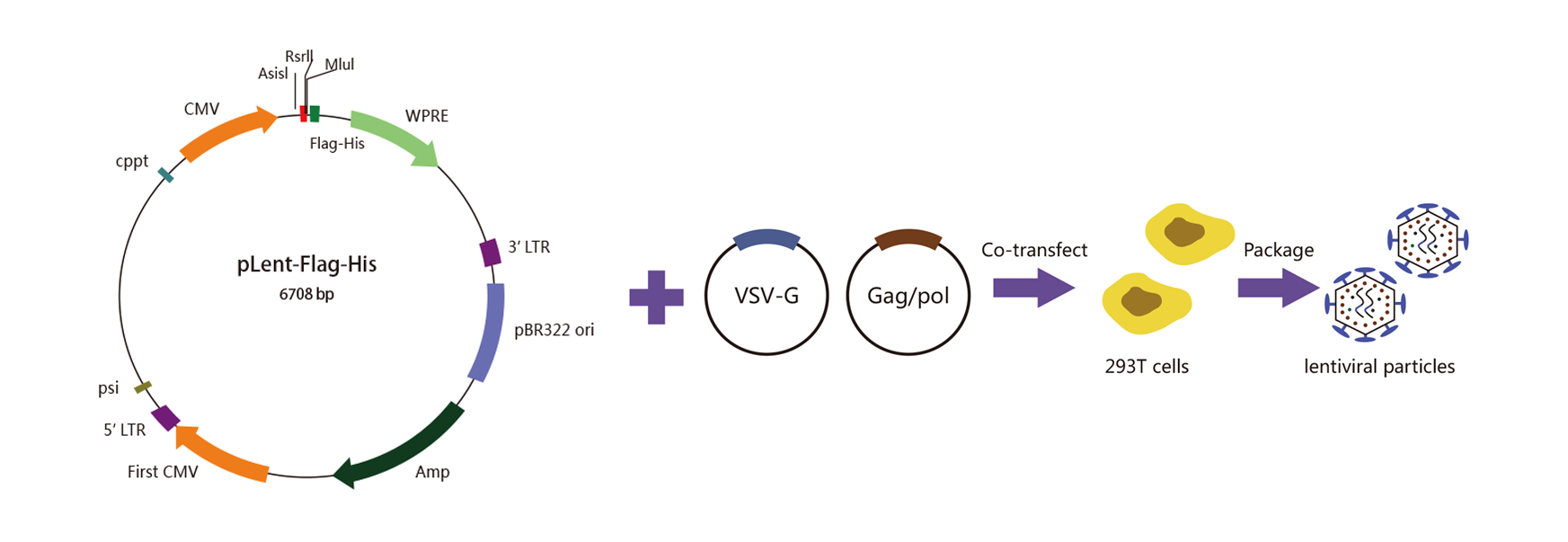
|
维真生物可以提供从LV质粒构建至病毒包装的所有服务外包项目。而且,维真生物全套的LV载体和表达系统可用于体内和体外表达human/mouse/rat ORFs,lncRNA,circRNA,shRNAs和CRISPR/gRNA。下面是LV从头合成的流程:
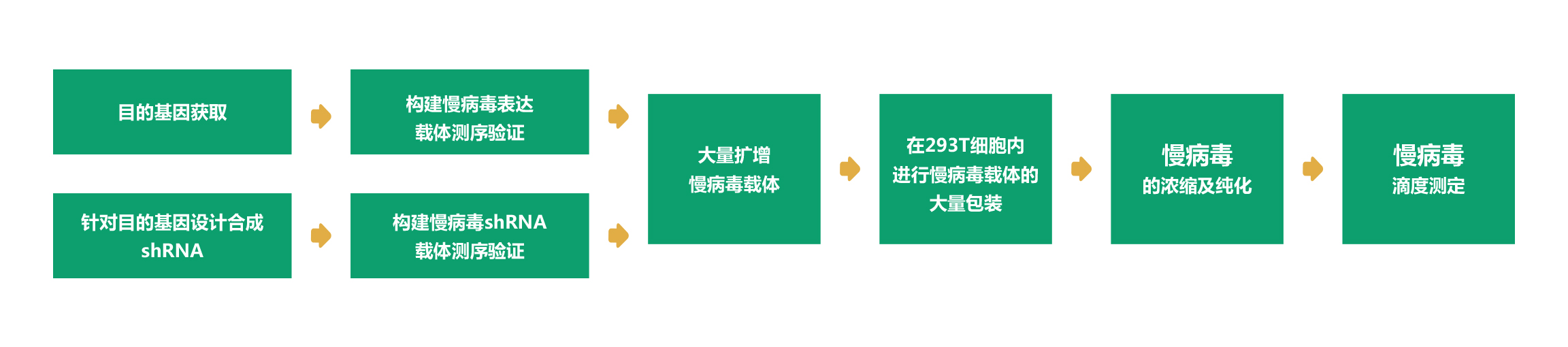
|
维真生物采用蔗糖密度梯度超速离心法对LV病毒进行纯化。它利用不同颗粒之间存在的沉降系数差,在一定离心力作用下,颗粒各自以一定速度沉降,在密度梯度不同区域上形成区带。下图是LV病毒纯化的示意图:
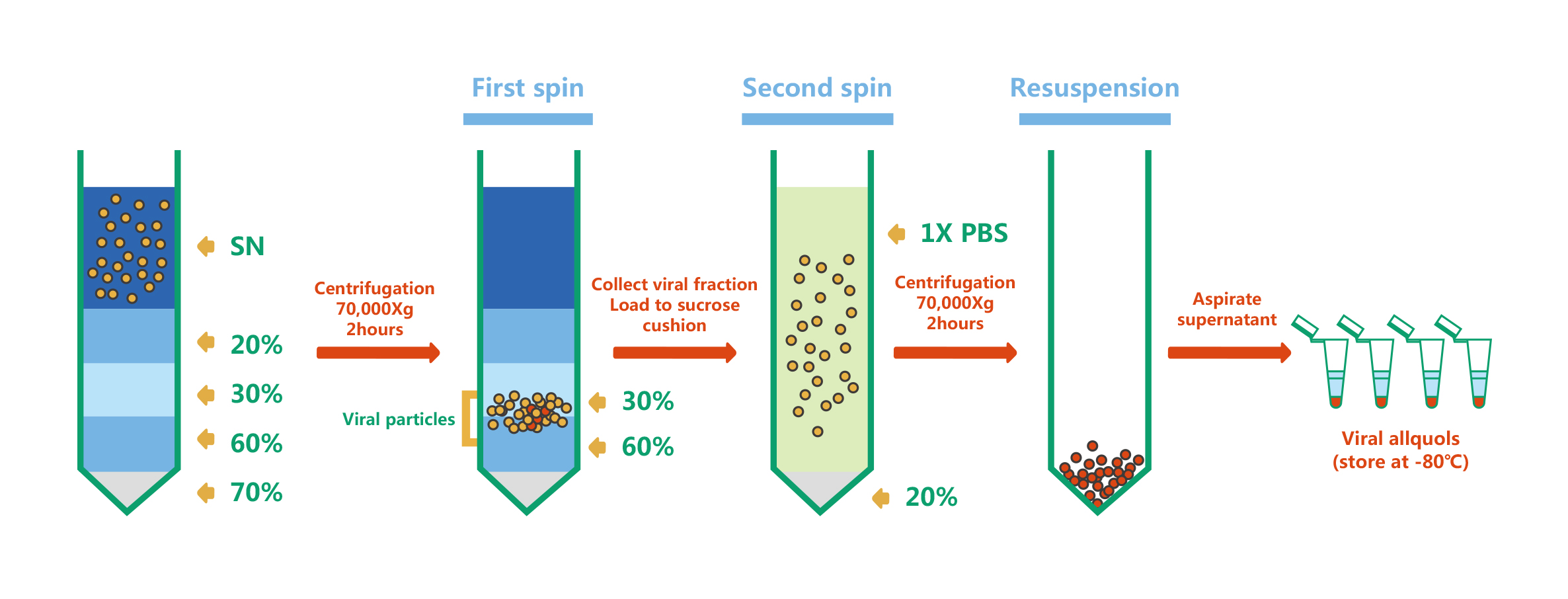
|
维真生物的LV病毒采用孔稀释法进行标定。
孔稀释法是通过数荧光的方法测定慢病毒滴度,得到的结果为有感染能力的慢病毒颗粒数。下图是维真某慢病毒(3.20×108TU/mL)孔稀释法测滴度时的荧光图:
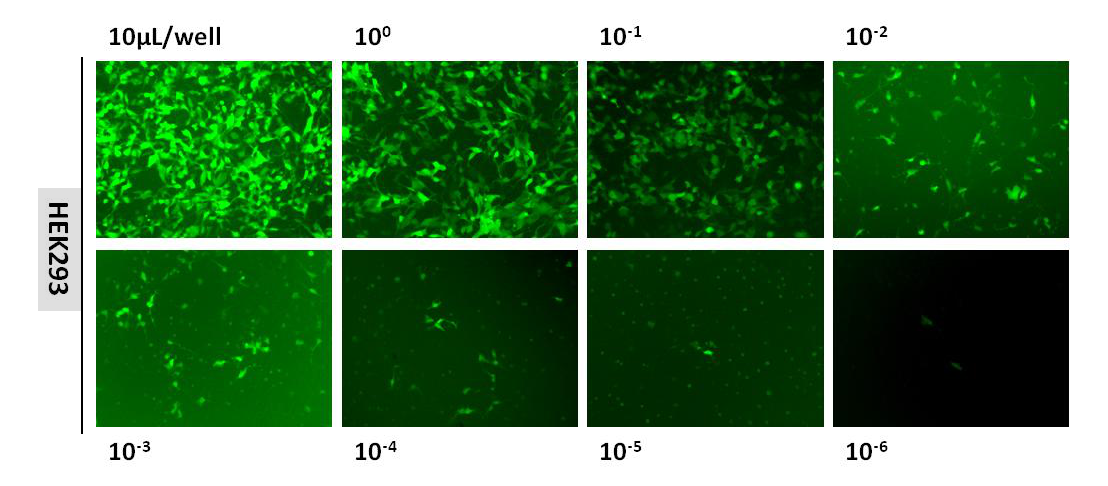
|
|
↑↑ 点击返回,查看其它部分 ↑↑ |
|
1. 载体全: |
多种表达载体、多种标签载体和多种报告基因可供客户选择。 |
|
2. 货期短: |
Human ORF cDNA克隆和Human microRNA克隆可在2-3天穿梭至慢病毒载体,个性化定做慢病毒; |
|
3. 服务优: |
可根据客户具体实验,由公司专业技术人员提供载体构建方案,提供特殊定制服务。 |
|
4. 赠送: |
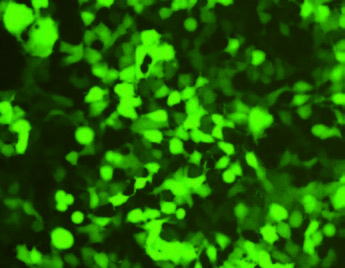
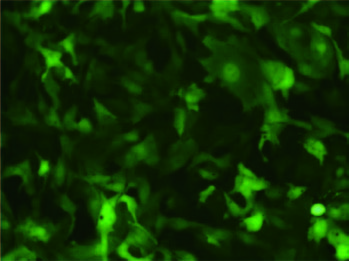
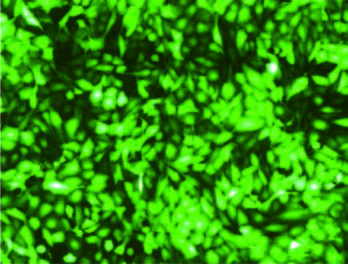
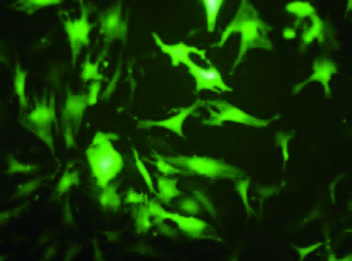
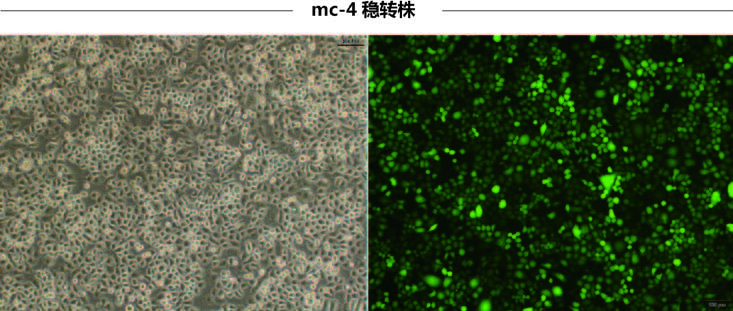
赠送慢病毒助感染试剂,这对于难感染细胞有很大的帮助,下图是助感染试剂在HepG2和CNE2细胞中的效果图。 |
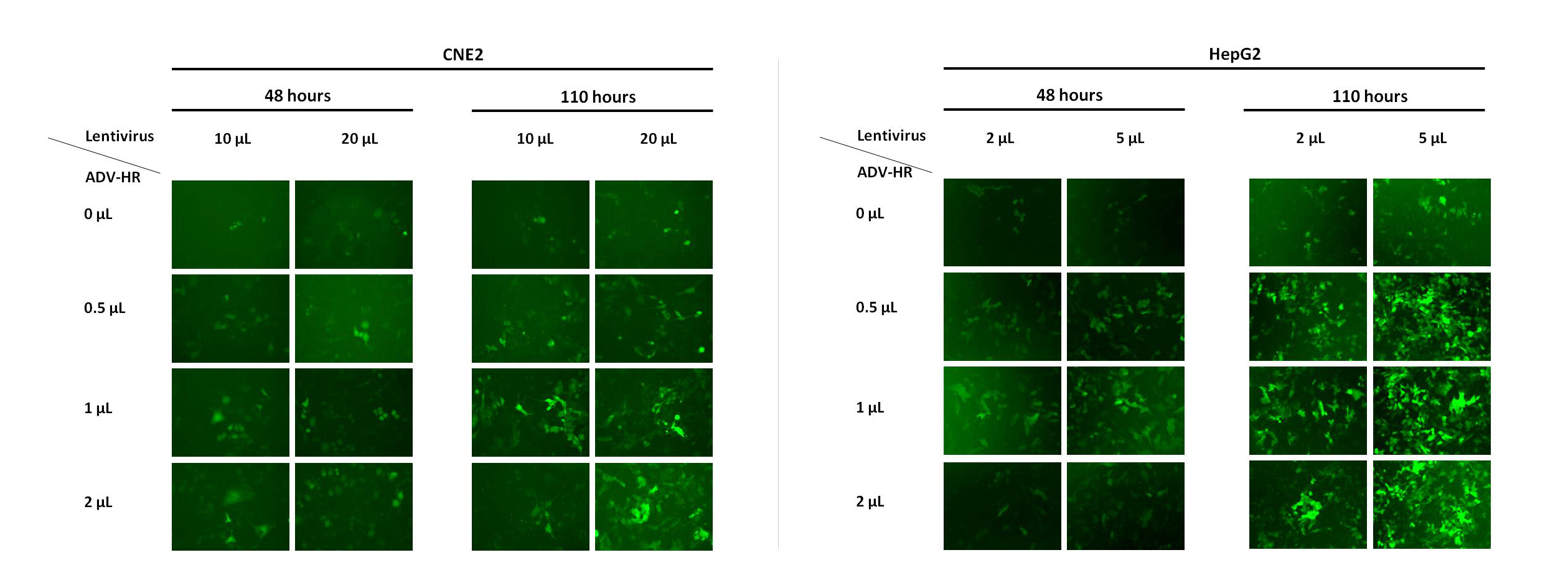
|
|
|
|
↑↑ 点击返回,查看其它部分 ↑↑ |
1、什么情况下选用慢病毒?
慢病毒的宿主范围比较广,既能感染分裂细胞,也能感染非分裂的细胞,在体内外研究中均可选择慢病毒作为病毒载体。此外,慢病毒也是构建稳转株的理想病毒。
2、慢病毒载体可插入多大的 ORF 片段?
一般情况下,慢病毒载体可容纳6 kb插入片段。然而,插入的ORF基因大于3 kb时会大大降低病毒的包装效率,进而影响病毒滴度甚至基因表达。
3、用于慢病毒感染的细胞接种量是多少?
将状态良好的目的细胞接种到24孔板,细胞浓度一般为1×105/mL细胞,接种细胞数量需要考虑到细胞活力因素、状态因素、生长快慢因素、分裂因素等,一般是保证病毒感染时细胞汇合率达到50%-70%为佳。
4、什么是 MOI ?
MOI (multiplicity of infection),即感染复数,指的是感染时病毒与细胞数量的比值。一般认为MOI是一个比值,没有单位。
5、慢病毒是否能够稳定表达基因?
是的。慢病毒能将外源基因整合到宿主基因组中,不会随着细胞分裂传代而丢失,因此能实现外源基因的长期稳定表达。
6、维真慢病毒的包装周期是多久?
如果客户提供构建好的慢病毒载体,那么包装时间一般为2周左右,若包含前期载体构建等的时间,时间大概在5周左右。
7、慢病毒感染细胞后,目的基因的表达什么时间达到峰值?
慢病毒感染目的细胞后,一般情况下,在72-96h目的基因的表达能达到峰值,但是对于一些特殊细胞、如增殖传代比较慢的细胞,蛋白表达达到峰值则需要更长的时间。
8、怎样提高慢病毒的感染效率?
细胞良好的生长状态和感染时适合的 MOI 值是达到高感染效率的保证。一般可以通过提高感染时的 MOI 值来提高病毒的感染效率,必要时可在感染时加入助感染试剂ADV-HR来提高慢病毒的感染效率。
9、慢病毒感染后,细胞状态很差,甚至出现死亡,为什么?
慢病毒会对细胞造成一定的毒性,不同细胞对毒性的耐受力不同。建议调整并降低感染时使用的MOI值,即可在细胞准备时增加铺板细胞的汇合率(可提高至 70%),调整感染时加入的病毒量。此外,还可选择在感染4小时后换液,用新鲜的完全培养液继续培养观察。
10、慢病毒、腺病毒、腺相关病毒三种病毒有什么区别?如何选择?
对于转染困难的细胞,科研工作者通常会选择病毒来导入目的基因。目前,腺病毒、慢病毒和腺相关病毒是常用的病毒载体工具。那么如何选择适合您实验体系的病毒工具,请参考下面3种病毒工具的比较:
|
病毒表达系统 |
腺病毒 |
腺相关病毒 |
慢病毒 |
|
病毒基因组 |
dsDNA |
ssDNA |
ssRNA |
|
病毒外壳 |
无 |
无 |
具有包膜蛋白 |
|
基因组大小 |
38-39kb |
5kb |
9kb |
|
包装容量 |
7.5kb |
4.7kb |
6kb |
|
感染的细胞类型 |
分裂细胞和非分裂细胞 |
分裂细胞和非分裂细胞 |
分裂细胞和非分裂细胞 |
|
整合至宿主基因组 |
非整合 |
非整合 |
整合 |
|
表达丰度 |
高水平表达 |
高水平表达 |
中到高水平表达 |
|
表达时间 |
快(1-2天) |
1-2周(体内) |
慢(2-4天) |
|
外源基因持续表达时间 |
短暂 |
潜在的持久 |
长久 |
|
免疫反应 |
较高 |
极低 |
低 |
|
相对病毒滴度 |
10E10pfu/mL |
10E13VG/mL |
10E8TU/mL |
|
生物安全等级 |
BSL-2 |
BSL-2 |
BSL-2 |
|
↑↑ 点击返回,查看其它部分 ↑↑ |

400-077-2566

service@wzbio.cn