当前位置:首页 > 新闻中心 > 新品发布
当前位置:首页 > 新闻中心 > 新品发布
1.研究背景
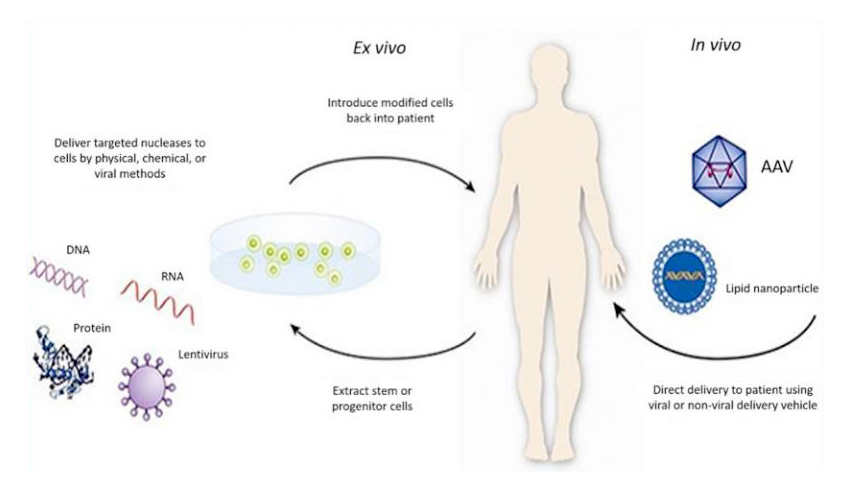
基因治疗是一种新型的治疗手段,它可以治疗多种疾病,包括癌症、遗传性疾病、神经性疾病、感染性疾病、心血管疾病和自身免疫性疾病等,它针对的是疾病的根源而不仅仅是疾病的症状。如此有效的治疗方法有什么特殊性呢?其实很简单,它就是将外源正常基因借助一定的技术或者载体导入靶细胞,从而纠正或补偿缺陷和异常基因引起的疾病,以达到治疗疾病的目的(图1)。
|
|
图1:基因治疗的示意图(来源于:https://www.fda.gov/)
|
|
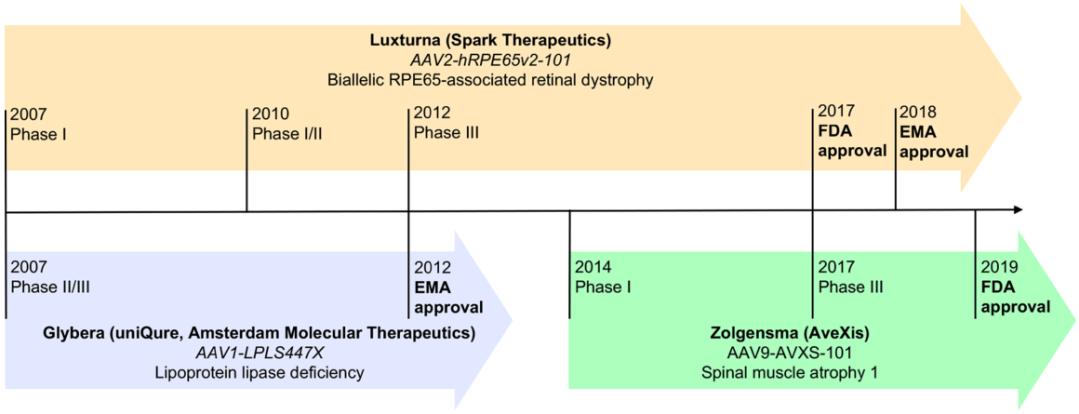
图2:3款获批上市的基于AAV载体的基因药物的时间表
(Lugin, M. L., et al. (2020). ACS Nano 14(11): 14262-14283.)
上述取得的成就,再加上美国临床试验数据库(ClinicalTrials.gov)中的一系列临床前和临床研究均证实了AAV载体的安全性和有效性。但是野生型AAV毕竟不是为了递送疾病治疗基因而进化的。而且,体内预存的中和抗体和AAV载体的承载能力进一步限制了AAV载体的广泛应用。目前,科学家们研究的重点主要集中在工程化AAV载体,用于提高其安全性、靶向性和转导效率。工程化AAV载体的构建方法多种多样,本篇我们将重点跟大家分享衣壳修饰的工程化AAV载体(图3)。
|
|
图3:工程化AAV载体的示意图
2.AAV基因组结构
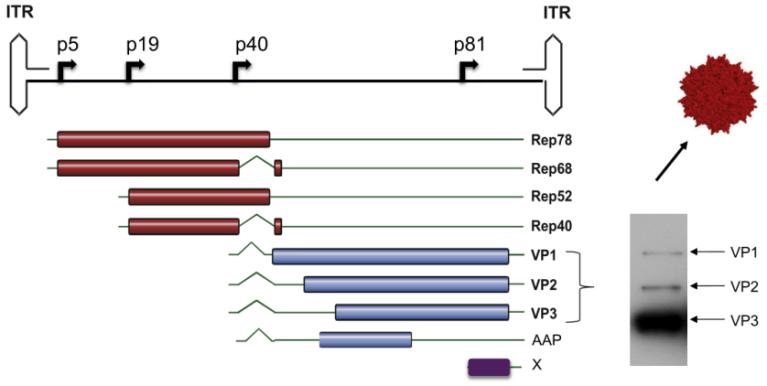
了解AAV基因组结构是构建工程化AAV的基础。野生型AAV含有约4.7kb的单链线性DNA基因组,三个基因Rep, Cap和X的两端各由145bp的ITR组成,可形成T型的发夹结构(图4)。Rep基因编码病毒转录调控、复制和包装必需的四个调控蛋白:Rep78, Rep68, Rep52 和Rep40。Cap基因编码三个重叠的结构蛋白(VP1, VP2, VP3,其表达比例为1:1:10)和VPs入核必需的装配激活蛋白AAP(assembly-activating protein)。X基因编码基因组复制中具有支持功能的蛋白质。
|
|
图4:AAV2载体的基因组结构
(Buning, H. and A. Srivastava (2019). Mol Ther Methods Clin Dev 12: 248-265.)
3.AAV衣壳结构
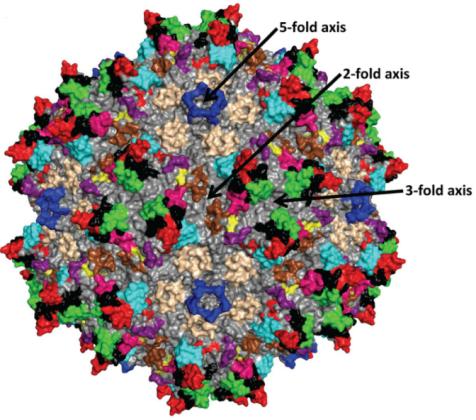
AAV衣壳结构的解析有助于理解AAV如何感染细胞和确定工程化AAV衣壳修饰的位置。腺相关病毒的衣壳呈T=1二十面体(二十面体是由20个三角形围成的凸多面体,每5个三角形围出一个五倍顶,通过每一对相对着的五倍顶有一个五倍旋转轴;通过每一对相对着的三角形中心有1个三倍旋转轴;通过每一对相对着的棱的中点有1个二倍旋转轴)。它由60个VP单体通过这些旋转轴的相互作用装配而成,这些VP单体可以全部为VP3,也可以由VP1、VP2、VP3共同构成。
AAV衣壳表面的显著特征是二倍凹地,三倍凸起和五倍通道(图5)。圆柱形通道连通衣壳内外,是AAV DNA进入衣壳的地方,它参与 Rep 蛋白结合、衣壳蛋白装配、VP1 N 末端外露、AAV病毒感染等多个过程;二倍凹地是病毒衣壳最薄的地方;三倍凸起的主要作用是识别受体。
|
|
图5:由60个VP3装配成的AAV2衣壳的拓扑结构
(Tseng, Y. S. and M. Agbandje-McKenna (2014). Front Immunol 5: 9.)
|
|
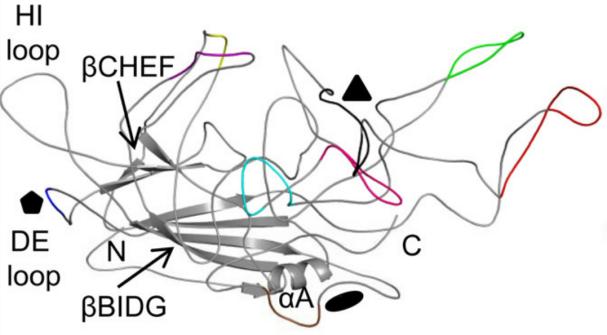
图6:AAV2 VP3单体的拓扑结构。9个VRs被用不同的颜色标记:
I,紫色;II,蓝色;III,黄色;IV,红色;V,黑色;VI,粉红色;VII,青色;VIII,绿色;IX,巧克力色
(Gurda, B. L., et al. (2013). J Virol 87(16): 9111-9124.)
4.衣壳修饰的工程化AAV
特定的VP亚基已被作为衣壳修饰的靶点,例如去除免疫原基序,整合标签或荧光基团和重新靶向等。
AAV2是研究相对成熟的血清型。以AAV2为骨架,在公共VP3区,大量研究表明,I-587(VP1氨基酸数)和I-588为较常用的衣壳修饰位置,因为这2个位置地处三倍凸起附近,可以接受长达34个氨基酸长度的肽链插入,而不影响衣壳化和基因组包装。而且,外源肽链的插入使得AAV2第一受体结合基序被修饰,最终赋予AAV2变体新的靶向性。Buning, H.等人汇总了在I-587和I-588处插入肽链的AAV2变体(表1)。维真生物现拥有表1所列的所有AAV Cap质粒,数量近70种,可直接进行AAV病毒包装。
包装表1所列AAV血清型,年中让利仅需8折,还送免费对照!
我们承诺:包装不成功不收费!
表1:在I-587和I-588处插入肽链的AAV2变体
| Serotype | Position | Name | Target Cell Type | Insert |
| AAV2 | I-587 | AAV-I-587 | β1-integrin positive tumor cells | QAGTFALRGDNPQG |
| AAV2 | I-587 | AAV-588NGR | CD13-positive tumor cells | NGRAHA |
| AAV2 | I-587 | AAV-MO7A | tumor cells | RGDAVGV |
| AAV2 | I-587 | AAV-MO7T | tumor cells | RGDTPTS |
| AAV2 | I-587 | AAV-MecA | tumor cells | GENQARS |
| AAV2 | I-587 | AAV-MecB | tumor cells | RSNAVVP |
| AAV2 | I-587 | rRGD587 | αv integrin positive tumor cells | CDCRGDCFC |
| AAV2 | I-587 | AAV-C4 | tumor cells | PRGTNGP |
| AAV2 | I-587 | AAV-D10 | tumor cells | SRGATTT |
| AAV2 | I-587 | AAV-SIG | endothelial cells | SIGYPLP |
| AAV2 | I-587 | AAV-MTP | endothelial cells | MTPFPTSNEANL |
| AAV2 | I-587 | AAV-QPE | endothelial cells | QPEHSST |
| AAV2 | I-587 | AAV-VNT | endothelial cells | VNTANST |
| AAV2 | I-587 | AAV-CNH | endothelial cells | CNHRYMQMC |
| AAV2 | I-587 | AAV-CAP | endothelial cells | CAPGPSKSG |
| AAV2 | I-587 | AAV-EYH | smooth muscle cells | EYHHYNK |
| AAV2 | I-587 | AAV587MTP | skeleton muscle cells | ASSLNIA |
| AAV2 | I-587 | AAV-r3.45 | neuronal stem cells | TQVGQKT |
| AAV2 | I-587 | AAV2-LSS | CNS | LPSSLQK |
| AAV2 | I-587 | AAV2-PPS | CNS | DSPAHPS |
| AAV2 | I-587 | AAV2-TLH | CNS | GWTLHNK |
| AAV2 | I-587 | AAV2-GMN | CNS | GMNAFRA |
| AAV2 | I-587 | AAV-Kera1 | keratinocytes | RGDTATL |
| AAV2 | I-587 | AAV-Kera2 | keratinocytes | PRGDLAP |
| AAV2 | I-587 | AAV-Kera3 | keratinocytes | RGDQQSL |
| AAV2 | I-587 | AAV-588Myc | none | EQLSISEEDL |
| AAV2 | I-587 | AAV2.N587_R588insBAP | adaptor | GLNDIFEAQKIEWHE |
| AAV2 | I-587 | AAV2Ald13 | adaptor | LCTPSRAALLTGR |
| AAV2 | I-587 | DMD4 | vaccine | QVSHWVSGLAEGSFG |
| AAV2 | I-587 | DMD6 | vaccine | LSHTSGRVEGSVSLL |
| AAV2 | I-588 | A588-RGD4C | av integrin-positive tumor cells | CDCRGDCFC |
| AAV2 | I-588 | A588-RGD4CGLS | av-integrin positive tumor cells | CDCRGDCFC |
| AAV2 | I-588 | AAV-VTAGRAP | tumor cells | VTAGRAP |
| AAV2 | I-588 | AAV-APVTRPA | tumor cells | APVTRPA |
| AAV2 | I-588 | AAV-DLSNLTR | tumor cells | DLSNLTR |
| AAV2 | I-588 | AAV-NQVGSWS | tumor cells | NQVGSWS |
| AAV2 | I-588 | AAV-EARVRPP | tumor cells | EARVRPP |
| AAV2 | I-588 | AAV-NSVSLYT | tumor cells (CML) | NSVSLYT |
| AAV2 | I-588 | AAV-LS1 | tumor cells (CML), CD34+cells | NDVRSAN* |
| AAV2 | I-588 | AAV-LS2 | tumor cells (CML), CD34+cells | NESRVLS |
| AAV2 | I-588 | AAV-LS3 | tumor cells (CML), CD34+cells | NRTWEQQ |
| AAV2 | I-588 | AAV-LS4 | tumor cells (CML), CD34+cells | NSVQSSW |
| AAV2 | I-588 | AAV-RGDLGLS | tumor cells | RGDLGLS |
| AAV2 | I-588 | AAV-RGDMSRE | tumor cells | RGDMSRE |
| AAV2 | I-588 | AAV-ESGLSQS | tumor cells | ESGLSQS |
| AAV2 | I-588 | AAV-EYRDSSG | tumor cells | EYRDSSG |
| AAV2 | I-588 | AAV-DLGSARA | tumor cells | DLGSARA |
| AAV2 | I-588 | AAV-GPQGKNS | tumor cells | GPQGKNS |
| AAV2 | I-588 | AAV-NSSRDLG | endothelial cells | NSSRDLG |
| AAV2 | I-588 | AAV-NDVRAVS | endothelial cells | NDVRAVS# |
| AAV2 | I-588 | AAV-PRSTSDP | lung (maybe endothelial cells) | PRSTSDP |
| AAV2 | I-588 | AAV-DIIRA | endothelial cells | DIIRA |
| AAV2 | I-588 | AAV-SYENV | endothelial cells | SYENVASRRPEG |
| AAV2 | I-588 | AAV-PENSV | endothelial cells | PENSVRRYGLEE |
| AAV2 | I-588 | AAV-LSLAS | endothelial cells | LSLASNRPTATS |
| AAV2 | I-588 | AAV-NDVWN | endothelial cells | NDVWNRDNSSKRGGTTEAS |
| AAV2 | I-588 | AAV-NRTYS | endothelial cells | NRTYSSTSNSTSRSEWDNS |
| AAV2 | I-588 | rAAV2-ESGHGYF | pulmonary endothelial cells | ESGHGYF |
| AAV2 | I-588 | AAV-GQHPRPG | cardiomyoblasts | GQHPRPG+ |
| AAV2 | I-588 | AAV-PSVSPRP | cardiomyoblasts | PSVSPRP |
| AAV2 | I-588 | AAV2-VNSTRLP | cardiomyoblasts | VNSTRLP |
| AAV2 | I-588 | AAV-LSPVR | cardiomyoblasts | LSPVRPG |
| AAV2 | I-588 | AAV-MSSDP | cardiomyoblasts | MSSDPRRPPRDG |
| AAV2 | I-588 | AAV-GARPS | cardiomyoblasts | GARPSEVTTRPG |
| AAV2 | I-588 | AAV-GNEVL | cardiomyoblasts | GNEVLGTKPRAP |
| AAV2 | I-588 | AAV-KMRPG | cardiomyoblasts | KMRPGAMGTTGEGTRVTRE |
| AAV2 | I-588 | AAV588MTP | skeleton muscle | ASSLNIA |
除了I-587和I-588外,I-453、I-520&I-584、I-584、I-585也已被成功用于开发新细胞靶向性工程化AAV(表2)。
表2:非I-587和I-588衣壳变体
| Serotype | Position | Name | Target Cell Type | Insert |
| AAV2 | I-453 | rRGD453ko | av integrin-positive tumor cells | CDCRGDCFC |
| AAV2 | I-453 | AAV-MNVRGDL | endothelial cells | MNVRGDL |
| AAV2 | I-453 | AAV-ENVRGDL | endothelial cells | ENVRGDL |
| AAV2 | I-520 and I-584 | A520/N584 (RGD) | av integrin-positive tumor cells | CDCRGDCFC |
| AAV2 | I-584 | A584-RGD4C | av integrin-positive tumor cells | CDCRGDCFC |
| AAV2 | I-584 | A584-RGD4CALS | av integrin-positive tumor cells | CDCRGDCFC |
| AAV2 | I-585 | AAV-∆IV-NGR | CD13-positive tumor cells | NGRAHA |
| AAV2 | I-585 | AAV-PTP | Plectin-positive tumor cells | KTLLPTP |
正如研究背景中所述,体内预存中和抗体是限制AAV广泛临床应用的一大障碍。血清学研究表明,大多数人都接触过野生型AAV,因此体内可能已经形成针对AAV的中和抗体。这些中和抗体可能会干扰AAV进入靶细胞、细胞内运输和细胞核内的解包装,从而阻止了转导。据报道,抗AAV抗体在人群中的流行率约为40-80%。其中,抗AAV2的中和抗体在人群中相对更流行。为了克服这个障碍,科学工作者以其他低免疫原性血清型为骨架,并参考AAV2的衣壳修饰位点,开发了一系列新的工程化AAV(表3)。
表3:非AAV2骨架的工程化AAV
| Serotype | Position | Name | Target Cell Type | Insert |
| AAV1 | I-590 | BAP-AAV1 | Scavidin displaying BT4C (rat glioma) | GLNDIFEAQKIEWHE |
| AAV1 | I-590 | BAP-AAV1 | endothelial cells | GLNDIFEAQKIEWHE plus CDCRGDCFC(RGD4C) |
| AAV1 | I-590 | AAV1-RGD | tumor cells, endothelial cells | CDCRGDCFC |
| AAV1 | I-590 | AAV1-RGD/BAP (90/10) (mosaic capsid) | tumor cells, endothelial cells | CDCRGDCFC and GLNDIFEAQKIEWHE |
| AAV1 | I-590 | Tet1c-AAV1 (mosaic capsid) | tetanus toxin GT1b receptor positive cells | HLNILSTLWKYR |
| AAV1 | I-590a | AAV1.9-3-SKAGRSP | fibroblast | SKAGRSP |
| AAV5 | I-575 | BAP-AAV4 | tumor cells | GLNDIFEAQKIEWHE |
| AAV6 | I-585 | AAV6-RGD | tumor cells | RGD |
| AAV6 | I-585 plus Y705-731F+T492V | AAV6-RGD-Y705-731F+T492V | tumor cells | RGD |
| AAV6 | I-585plus Y705-731F+T492V+K531E | AAV6-RGD-Y705-731F+T492V+K531E | tumor cells | RGD |
| AAV8 | I-585c | AAV2/8-BP2 | on-bipolar cells | PERTAMSLP |
| AAV8 | I-590 | AAV8-ESGLSOS | tumor cells | ESGLSOS135 |
| AAV8 | I-590 | AAV8-ASSLNIA | heart (weakly improved transduction) | ASSLNIA122 |
| AAV8 | I-590d | AAV8-GQHPRPG | heart (weakly improved transduction) | GQHPRPG86 |
| AAV8 | I-590d | AAV8-SEGLKNL | liver | SEGLKNL |
| AAV9 | I-589 | AAV-SLRSPPS | endothelial cells, smooth muscle cells | SLRSPPS |
| AAV9 | I-589 | AAV-RGDLRVS | endothelial cells, smooth muscle cells | RGDLRVS |
| AAV9 | I-589d | AAV9-NDVRAVS | endothelial cells | NDVRAVS82 |
| AAV9 | I-589d | AAV9-ESGLSOS | tumor cells (weak targeting) | ESGLSOS135 |
| AAV9 | I-588 | AAV-PHP.B | CNS | TLAVPFK |
| AAV9 | I-588 | AAV-PHP.A | CNS | YTLSQGW |
| AAV9 | I-588 | AAV9-7m8 | retinal cells | LGETTRP80 |
| AAV9P1 | not disclosed | AAV9P1 | neuronal progenitor cells | RGDLGLS |
如若了解靶向同一类细胞的所有血清型间的效果差异,请参考Reference 2中对应的参考文献。

|
维真提供快速即用工程化AAV——保证包装成功! |
|
您可以从表1中选择您感兴趣的AAV Cap质粒,我们将1-2周交付您即用型AAV病毒,并且承诺包装不成功不收费,大大加快您的科研进程! 此外,如果您对表2和表3中的质粒感兴趣,我们也可以帮您进行个性化定制,保证包装成功! |
|
咨询订购:①拨打热线电话400-077-2566; ②致电您的区域经理;③添加维真客服咨询! |
目前AAV基因药物的临床研究大多数采用的是天然的衣壳,这些衣壳极易成为宿主酶的作用靶点,从而影响它们的整体性能。此外,天然AAV血清型具有组织亲噬性,但通常趋向性比较广泛。因此,必须使用高剂量的病毒才能达到治疗效果。而且,AAV载体的包装容量和大多数人群中已存的中和抗体进一步限制了AAV的临床应用。为了克服这些限制,科学工作者将研究的重点聚焦在工程化AAV,以进一步提高其安全性、有效性和特异性,从而推动基因治疗领域的快速发展。在当今基因治疗的研究热潮下,为了加快您的科研进程,我们建立了工程化AAV库,旨在为基因治疗的发展贡献自己的一份力。
6.参考文献
1. Lugin, M. L., et al. (2020). "Synthetically Engineered Adeno-Associated Virus for Efficient, Safe, and Versatile Gene Therapy Applications." ACS Nano 14(11): 14262-14283.
2. Buning, H. and A. Srivastava (2019). "Capsid Modifications for Targeting and Improving the Efficacy of AAV Vectors." Mol Ther Methods Clin Dev 12: 248-265.
3. Pipe, S., et al. (2019). "Clinical Considerations for Capsid Choice in the Development of Liver-Targeted AAV-Based Gene Transfer." Mol Ther Methods Clin Dev 15: 170-178.
4. Tseng, Y. S. and M. Agbandje-McKenna (2014). "Mapping the AAV Capsid Host Antibody Response toward the Development of Second Generation Gene Delivery Vectors." Front Immunol 5: 9.
5. Gurda, B. L., et al. (2013). "Capsid antibodies to different adeno-associated virus serotypes bind common regions." J Virol 87(16): 9111-9124.

400-077-2566

service@wzbio.cn