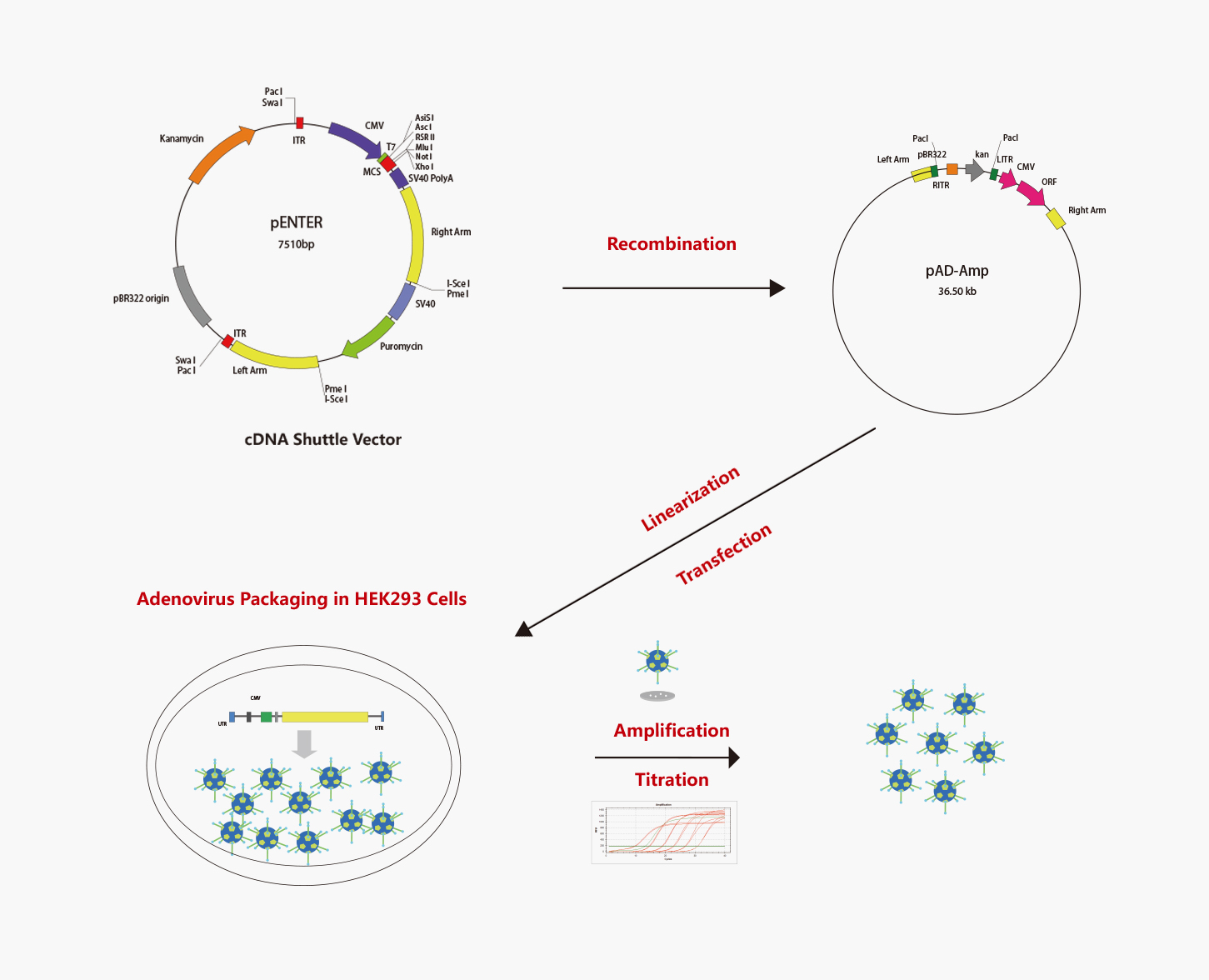
腺病毒(Adenovirus,Ad)是一种无包膜的DNA病毒,其复制不依赖于宿主细胞的分裂。腺病毒拥有近50个血清型,大多数腺病毒载体都是基于血清型2和血清型5,通过转基因的方式取代E1和E3基因,降低病毒的复制能力。这些重组病毒仅在高水平表达E1基因的细胞中复制,因此是一种适用于治疗的高效控制系统。重组腺病毒是进行基因转移和表达的工具,功能强大并易于操作。腺病毒独特的生物学特征使它成为“载体的首选”,被科研工作者广泛使用。
维真生物成立之初,其研发团队就建成了涵盖12000多种人源基因的腺病毒库,具体信息见人源ORF腺病毒。产品自2013年面市后,公司也定制了很多腺病毒包装项目。目前,维真生物拥有十余年的腺病毒包装经验,提供从腺病毒质粒设计、构建、包装到表达分析的所有服务外包项目。全套的腺病毒载体和表达系统可用于体内外表达多物种ORF、lncRNA、circRNA、shRNA 和CRISPR/gRNA,可满足客户多样化的实验需求。

下面是具体腺病毒服务内容(具体规格、费用、货期可详询客服)
| 服务编号 | 服务类型 | 规格 | 目录价 | 货期 |
|---|---|---|---|---|
| WZ050001 | 小包装、非纯化 | 滴度=1×10E10-11 vp/ml;体积1ml | 询价 | 详询 |
| WZ050002 | 中包装、纯化 | 滴度≥1×10E10 pfu/ml;体积1ml | 询价 | 详询 |
| WZ050003 | 大包装、纯化 | 滴度≥1×10E11 pfu/ml;体积1ml | 询价 | 详询 |
*如您对上述服务感兴趣,请您点击“欢迎垂询”留下您的信息,我们将为您提供具体实验方案和项目价格。
|
↓↓ 分以下几个部分做详细介绍 ↓↓ |
腺病毒是一种直径为90-100nm的病毒颗粒,衣壳呈廿面体,由240个六邻体和12个五邻体组成。以五邻体蛋白为基底由衣壳表面伸出12根纤毛,纤毛顶端形成头节区。五邻体和纤毛的头节区可与细胞表面的受体结合,在病毒感染细胞过程中起着非常重要的作用。右面是腺病毒的结构示意图:

腺病毒是一种双链线性的DNA病毒,其基因组长度约为36kb。基因组两端各有一个103 bp的反向末端重复序列(inverted terminal repeat, ITR),参与病毒DNA的复制;ITR的内侧为病毒包装信号Ψ,参与腺病毒基因组的衣壳化。基因组包含早期表达的与腺病毒复制相关的E1~E4基因和晚期表达的与腺病毒颗粒组装相关的L1~L5基因。下图是人类血清5型腺病毒基因组的示意图:

基于人血清5型腺病毒的基因组结构,结合各基因的功能,科学家们开发了缺失E1和E3基因的Ad5腺病毒载体。E1基因在组装感染性病毒颗粒时必不可少,但是可以在HEK293包装细胞中得到补充,而E3基因不影病毒的包装。由于E1和E3基因的缺失,腺病毒载体可插入高达7.5kb的外源基因。
腺病毒感染细胞的第一步是通过纤突蛋白与细胞表面CAR结合。腺病毒粘附后,五邻体蛋白上的精氨酸-谷氨酸-天冬氨酸(RGD)与细胞表面的整合素αvβ3和αvβ5结合并相互作用使腺病毒进入细胞。下图是腺病毒感染细胞的示意图:

1、腺病毒感染的宿主细胞范围广,包括分裂细胞和不分裂细胞(某些抗腺病毒感染的淋巴瘤细胞除外)。
2、腺病毒感染效率高,在最佳感染条件下,感染率可达100%。
3、腺病毒的病毒滴度可以很高,纯化、浓缩后可达1012-13vp/ml(即1010-11pfu/ml)。
4、腺病毒易于扩增,而其他病毒比如慢病毒和腺相关病毒需要重新包装。
5、不整合到染色体中,无插入致突变性。
6、腺病毒理化性质稳定,4℃:数周。-80 ℃:数年。
Ad5腺病毒的高效感染依赖于靶细胞膜上的CAR和αvβ整合素,但是一些免疫细胞的细胞膜上却缺乏这些受体,如造血干细胞。这就使得研究人员寻找新的腺病毒血清型,最终发现F35血清型对造血干细胞有很强的靶向性,因此,嵌合型Ad5/F35应用而生。下面是Ad5 & Ad5/F35的结构示意图和区别:

| 腺病毒种类 | 载体特点 | 功能 | 感染细胞受体 | 包装系统 |
|---|---|---|---|---|
| AdV5 | 缺失E1和E3基因,允许插入长约8kb的外源基因 |
除免疫、血液类细胞外, 感染大部分细胞 |
CAR | AdMax |
| AdV5/F35 | 一个嵌合型腺病毒载体,主要结构是在AdV5的基础上,其受体结合位置的纤突(Fiber)改造成了F35型腺病毒(F35)的纤突(纤毛蛋白的Knob和shaft)。其细胞受体从AdV5的CAR变成了CD46,其他的基因仍旧保留AdV5载体的特性,能有效转导CAR表达不足的多种重要靶细胞,尤其是对肿瘤细胞及造型干细胞、间充质干细胞等有较高的感染效率。 | 理论上感染所有有细胞核的细胞 | CD46 | AdMax |
腺病毒应用策略及客户案例分享 |
|||||||||||||||||||||||
1、腺病毒体外应用实例(部分) |
|||||||||||||||||||||||
1.1 腺病毒感染人脐静脉内皮细胞HUVECs主要应用:利用腺病毒在HUVECs 中过表达野生型FGFR1 和Y463F、Y585F、Y653F 突变体,结果发现 VEGF-B 在过表达FGFR1 WT 的HUVECs 中增加了VEGFR1/FGFR1 复合物的形成,但在过表达FGFR1 Y463F、FGFR1 Y585F 或FGFR1 Y653F 突变体的HUVECs 中没有增加,表明酪氨酸残基Y463、Y585 和 Y653 在介导VEGF-B 诱导的VEGFR1/FGFR1 复合物形成中发挥作用。本研究揭示了VEGF-B 作为FGF2/ FGFR1 途径内源性抑制剂的新功能。
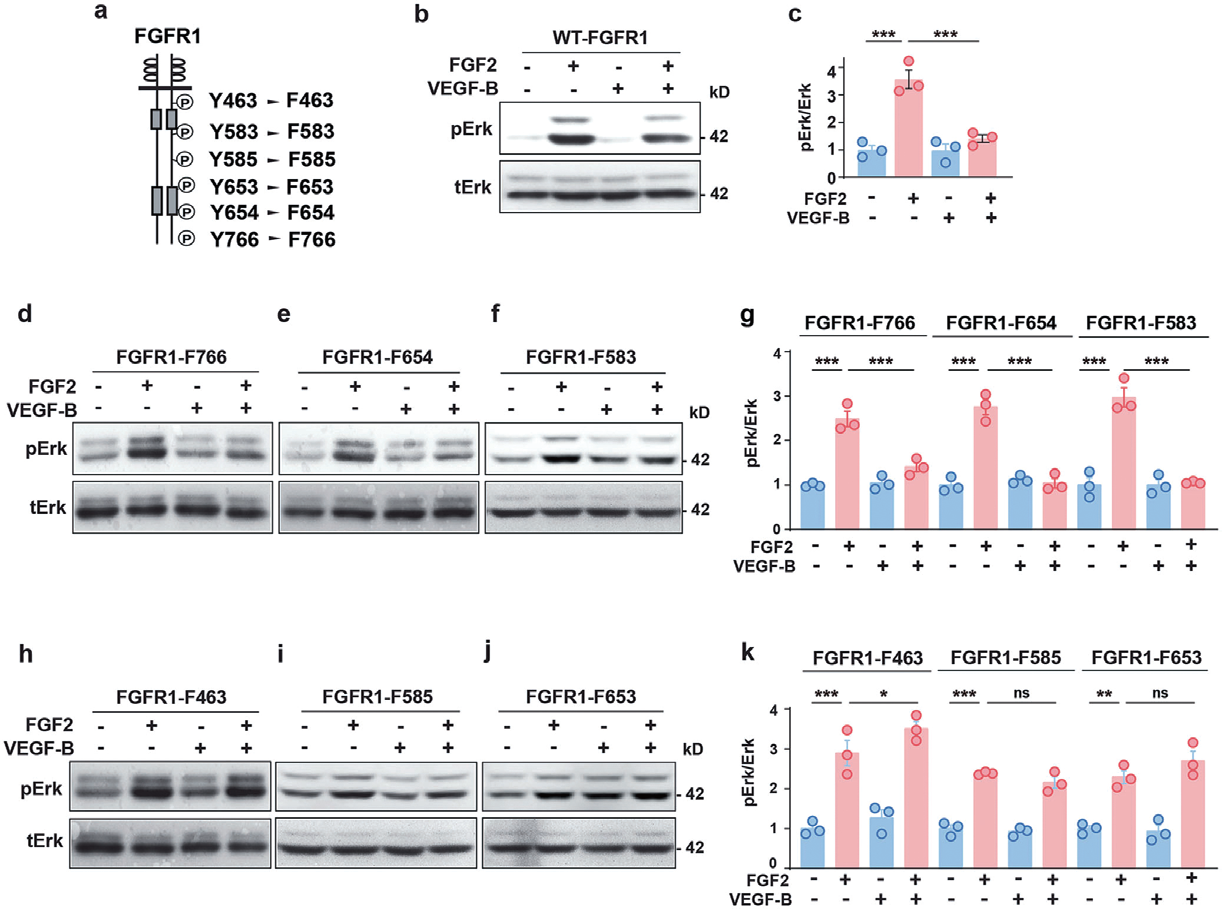
▲ VEGF-B 抑制FGF2 诱导的Erk 活化 |
1.2 腺病毒感染MIN6细胞和Islets细胞主要应用:胰腺β 细胞在维持胰岛素分泌和葡萄糖稳态中起着至关重要的作用,β 细胞质量和胰岛素分泌 不足是2 型糖尿病(T2D) 发展的必要条件。作者利用腺病毒进行Tipe1 拯救实验发现Tipe1 过表达可以 逆转其缺失引起的糖尿病表型,证实Tipe1 以细胞自主的方式促进β 细胞增殖和胰岛素分泌;为了确认 Gαs 是否与Tipe1 介导的胰岛β 细胞调控有关,将过表达Gnas 的腺病毒转导Tipe1 沉默的MIN6 细胞 和Ins2-Tipe1BKO 小鼠的胰岛,发现Gnas 的过表达逆转了MIN6 细胞的增殖,表明Gαs 参与Tipe1 介 导的β 细胞调控。
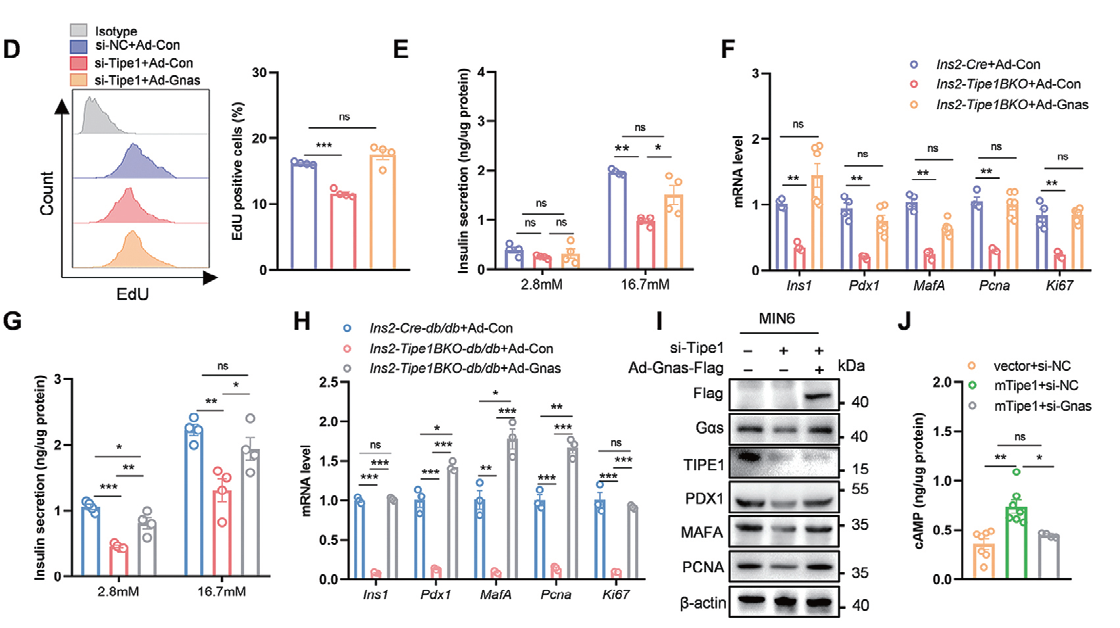
▲ Tipe1 通过Gαs/cAMP 通路调控β 细胞功能 |
||||||||||||||||||||||
1.3 自噬双标腺病毒感染人肝细胞主要应用:利用腺病毒介导的mCherry-GFP-LC3B 自噬双标探针感染人肝细胞HL-7702,研究PLK1 和 自噬间的关系。PLK1 的下调逆转了吉非替尼诱导的自噬通量,而PLK1 的过表达则增强了自噬通量。本 研究揭示了吉非替尼- 肝毒性途径,其中自噬通过COX6A1 降解促进细胞凋亡,并强调PLK1 的药理抑制 是提高吉非替尼癌症治疗安全性的一种有吸引力的治疗方法。
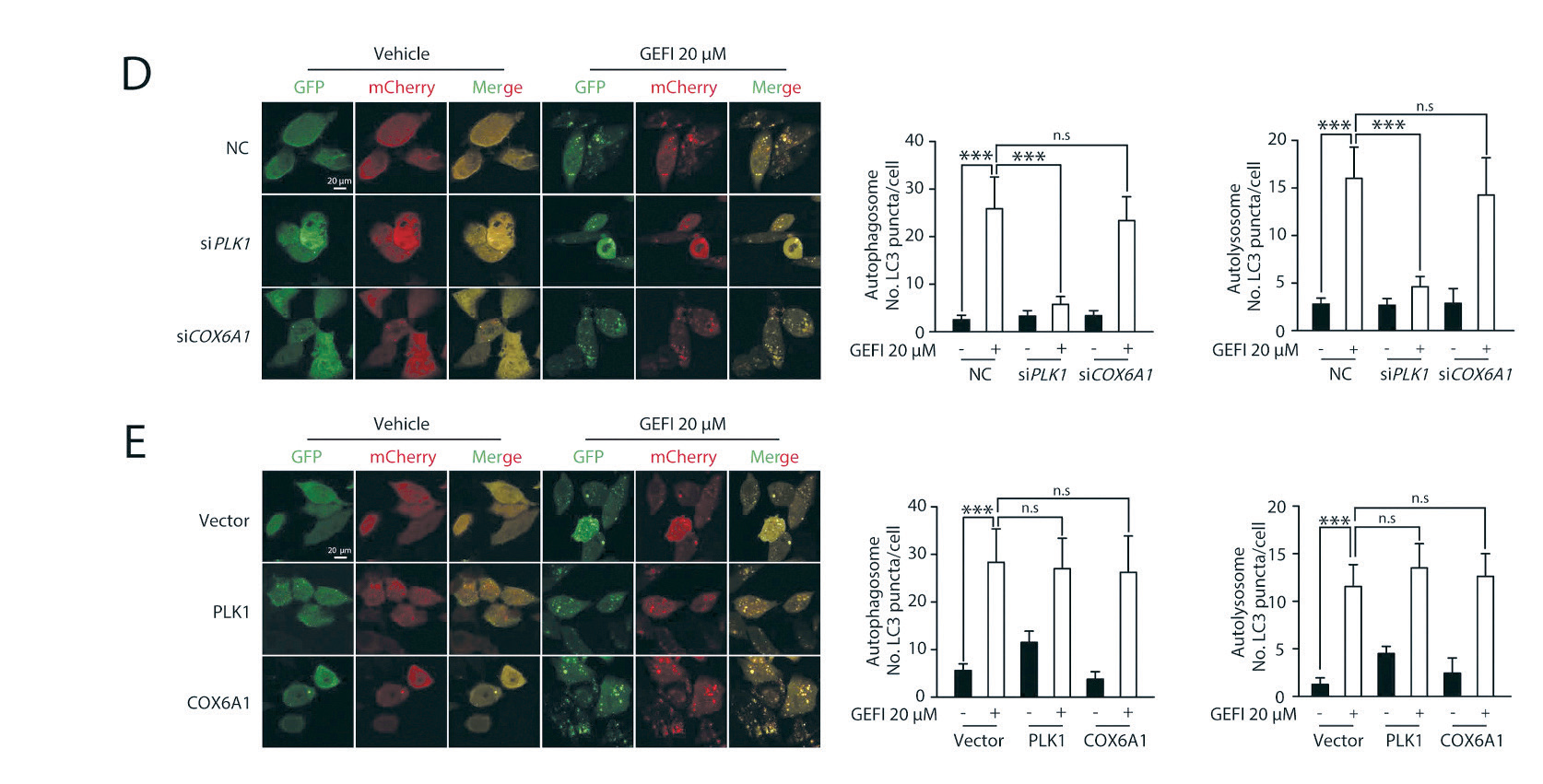
▲ PLK1 依赖性自噬参与吉非替尼诱导的肝毒性 |
1.4 腺病毒感染人肺癌细胞和人支气管上皮细胞主要应用:2019 冠状病毒病(COVID-19) 的发病机制受活性氧(ROS) 的影响,然而其具体机制尚不明确。 冠状病毒SARS-CoV-2 核衣壳(N)蛋白(S2NP)是该疾病的主要驱动因子,为了探究S2NP 是否影响 线粒体ROS 水平,作者利用腺病毒5 型载体在A549 细胞中过表达S2NP,发现S2NP 过表达组比对照 组表现出更高水平的线粒体ROS 水平,RNA 测序及ATP 含量测定表明S2NP 的表达阻碍了ATP 的产生, 进一步研究发现S2NP 的表达增强了复合物I 和III 的活性。
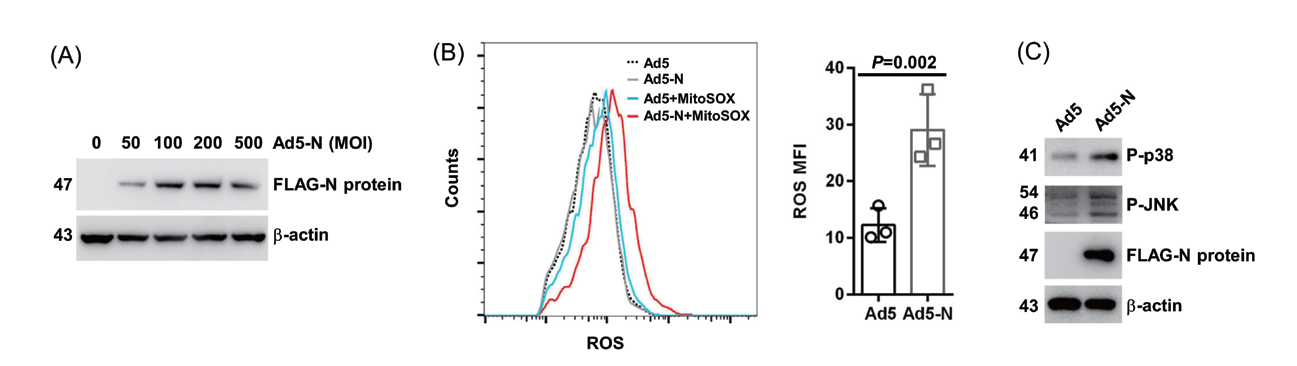
▲ S2NP 可提高线粒体ROS 水平 |
||||||||||||||||||||||
1.5 腺病毒感染新生大鼠心室肌细胞主要应用:利用腺病毒介导新生大鼠心室肌细胞(NRVMs) 中Prdm16 的过表达,显著增加了PPARα 和 PGC-1α 的反式激活;腺病毒介导的PPARα 和PGC-1α 过表达分别改善了PA 诱导的细胞凋亡和线粒体 功能障碍以及脂质摄取减少。本研究揭示PRDM16 以组蛋白赖氨酸甲基转移酶活性依赖的方式通过调节 PPARα 和PGC-1α,对T2DM 患者心肌脂质代谢和线粒体功能发挥保护作用。
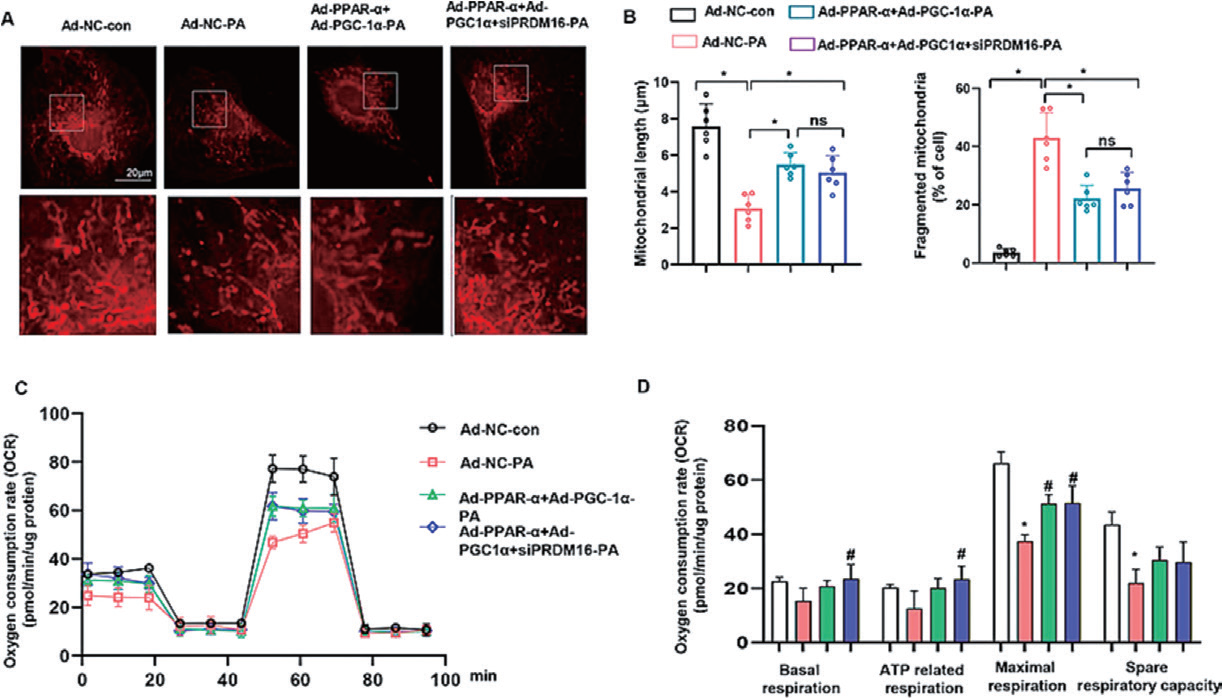
▲ PPARα 和PGC-1α 的过表达可逆转高脂环境下Prdm16 缺陷诱导的细胞功能障碍 |
1.6 腺病毒感染人骨肉瘤细胞主要应用:将Nrf2-shRNA 重组腺病毒载体转染至HOS 细胞中敲低Nrf2,当Fe3O4 浓度超过10μg Fe/ mL 时,转染shRNA-Nrf2 的癌细胞毒性逐渐增强,且Nrf2 的表达水平在0 ~ 10μg Fe/mL 范围内呈浓 度依赖性,表明Fe3O4 纳米颗粒处理后,癌细胞可以通过上调Nrf2 相关抗氧化系统表达来避免致死性铁 死亡。本研究为Fe3O4 纳米颗粒诱导铁死亡提供了新的机制见解和实用策略,合成的仿生纳米颗粒对骨肉 瘤表现出协同的铁死亡/ 免疫治疗作用。
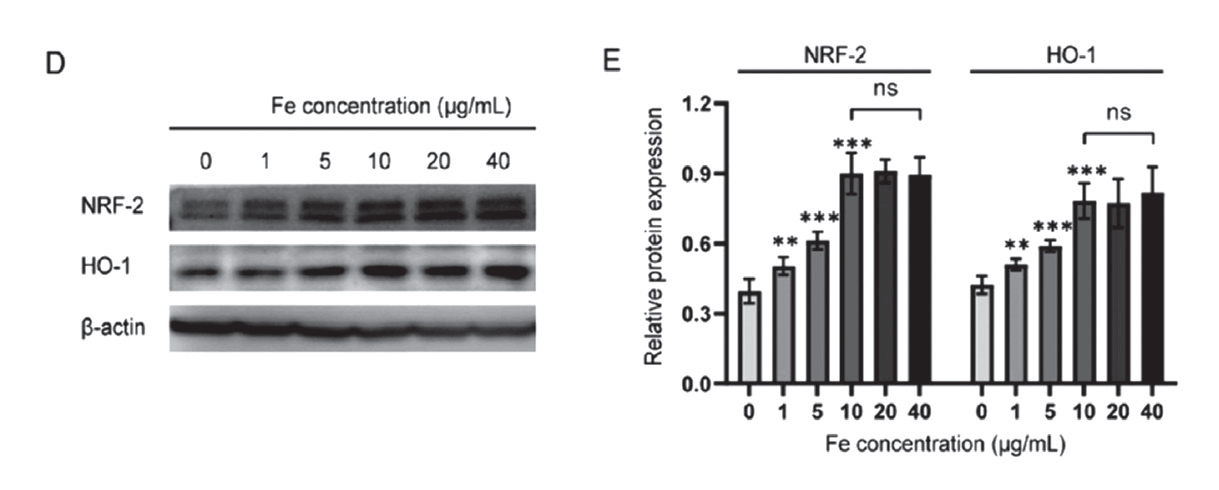
▲ 癌细胞通过激活Nrf-2 相关抗氧化系统来阻止Fe3O4 纳米颗粒诱导的致死性铁死亡 |
||||||||||||||||||||||
1.7 腺病毒感染小鼠骨髓巨噬细胞主要应用:用腺病毒shRNA 载体分别敲低Cytohesin-1/2/3 三种胞黏素的表达,然后诱导OC 分化。免疫 印迹实验证实每种胞黏素的敲除效率均在50% 以上,而不影响其他两种胞黏素含量,TRAP 染色发现OC 的 分化只受到Cytohesin-2 的影响,而Cytohesin-1 和Cytohesin-3 的下调不影响OC 的分化。本研究揭示 了Cytohesin-2 在调控破骨细胞分化和骨质疏松发展中的作用,为骨质疏松症的治疗提供了新的策略。
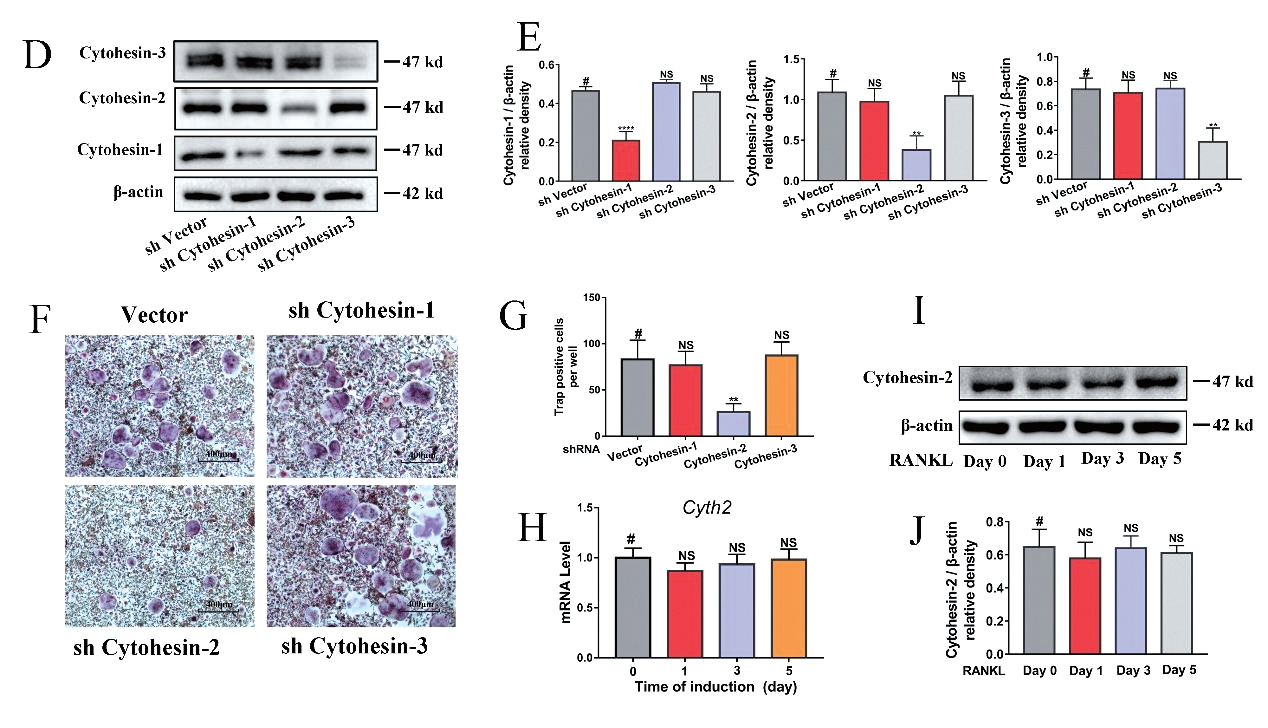
▲ SecinH3 通过抑制Cytohesin-2 来抑制OC 分化 |
1.8 腺病毒感染小鼠小胶质细胞主要应用:利用腺病毒在小胶质细胞中过表达和敲低SIRT5,发现在OGD/R 条件下,SIRT5 过表达显著 上调Il-1β、Il6、Tnf-α、Cxcl1 和Ccl2 等促炎因子和促炎介质iNOS 和Iba-1 的表达,SIRT5 的抑制则表 现出相反的效果,表明SIRT5 加重了小胶质细胞诱导的炎症反应。本研究揭示靶向SIRT5/ANXA1 可能是 一种用于缺血性中风和其他神经炎症性疾病的潜在治疗策略。
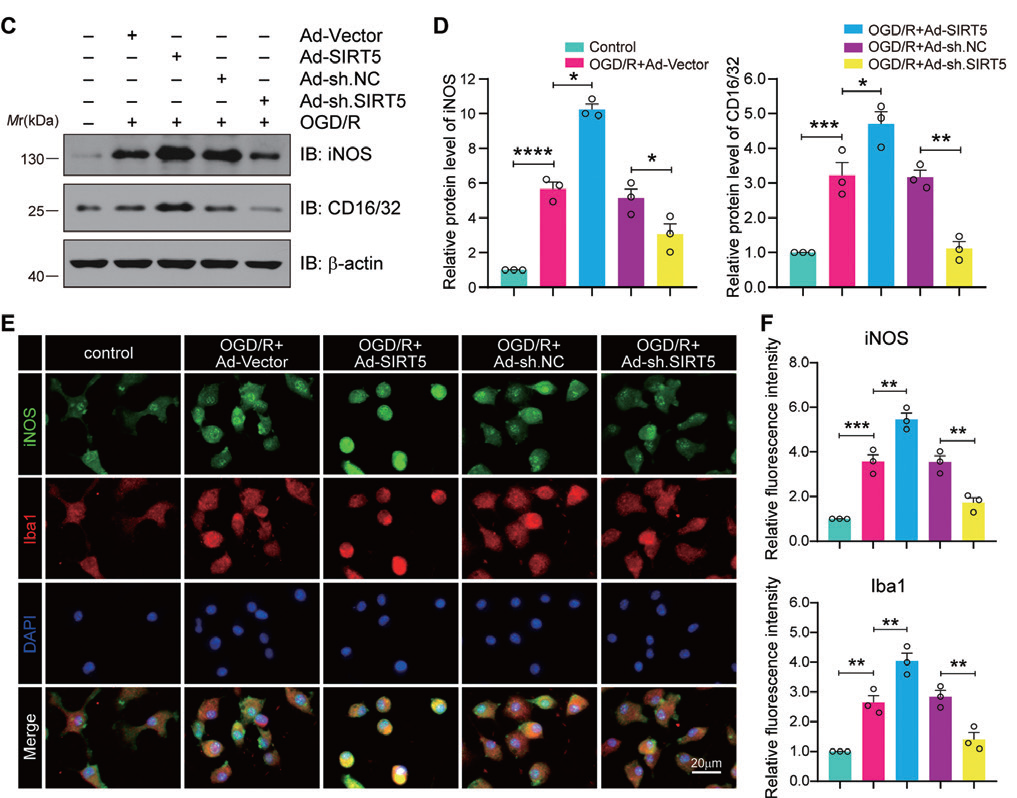
▲ SIRT5 加重小胶质细胞诱导的炎症反应 |
||||||||||||||||||||||
1.9 腺病毒感染ESC细胞、iPSC细胞主要应用:利用腺病毒载体过表达FAK 突变体(FAK Y397D,Tyr397 突变为Asp397),发现SAR 处理 后FOXA2 和SOX17 蛋白的显著上调,但FAK Y397D 过表达消除了SAR 的促进作用,表明FAK 磷酸化 在协调SAR 促进的DE 分化过程中发挥重要的调节作用。本研究揭示了SAR 通过抑制FAK-YAP 信号促进 内胚层分化。
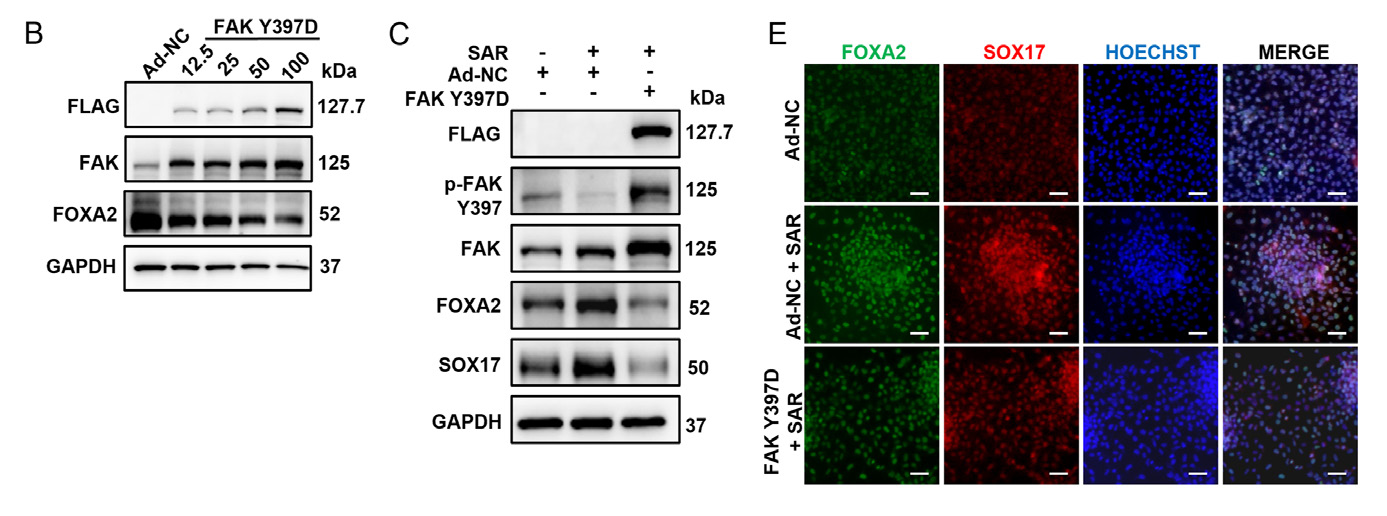
▲ SecinH3 通过抑制Cytohesin-2 来抑制OC 分化 |
1.10 腺病毒感染人牙龈成纤维细胞主要应用:作者利用Ad-CMV-mCherry-EGFP-LC3B 评估了牙龈卟啉单胞菌感染的HGFs 中的自噬水平: ebselen 单独处理对细胞自噬作用不大,但AgNPs 单独处理或AgNPs 和ebselen 联合处理均显著增加 了牙龈卟啉单胞菌感染的HGFs 中的自噬水平,并且ebselen 通过抑制NF-κB 和丝裂原活化蛋白激酶信 号通路的激活而发挥抗炎作用。证实协同抗炎作用归因于AgNPs 促进有害因子的清除和ebselen 抑制炎 症因子的表达。
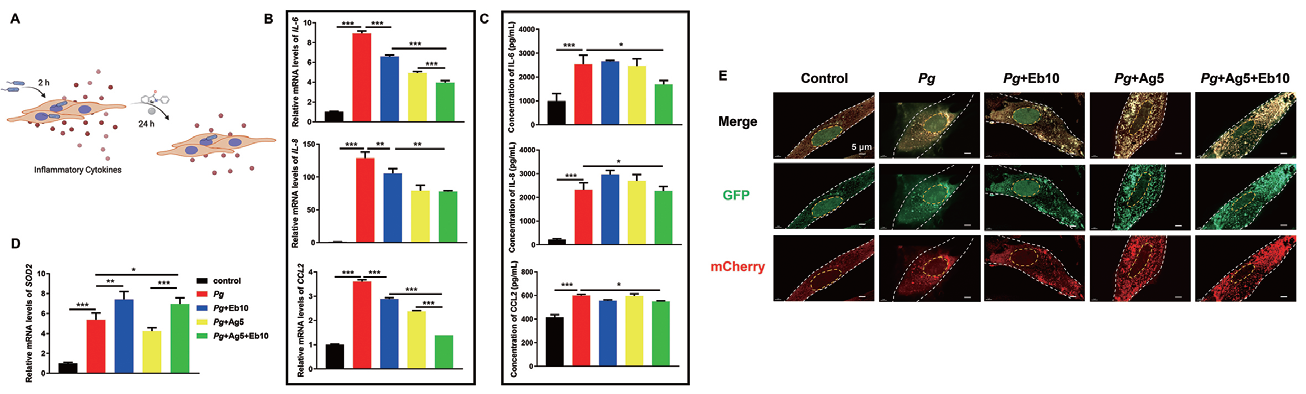
▲ AgNPs 和ebselen 的抗炎作用 |
||||||||||||||||||||||
1.11 腺病毒感染小鼠成肌细胞主要应用: 利用腺病毒介导的NFATc2 的过表达几乎完全逆转了Ezrin 敲低引起的MyHC 阳性多核肌管数 量的减少;腺病毒介导的NFATc4 的下调显著消除Ezrin 敲低对成肌细胞分化和融合的抑制作用。此外, NFATc2 的过表达或NFATc4 的下调均明显消除了Ad-shEzrin 对三核以下细胞和三核以上肌管中MyoG+ 和MEF2C+ 核数量和百分比的抑制作用。因此,Ezrin 通过NFATc2/c4-MyoG/MEF2C 参与成肌细胞的分 化和融合。
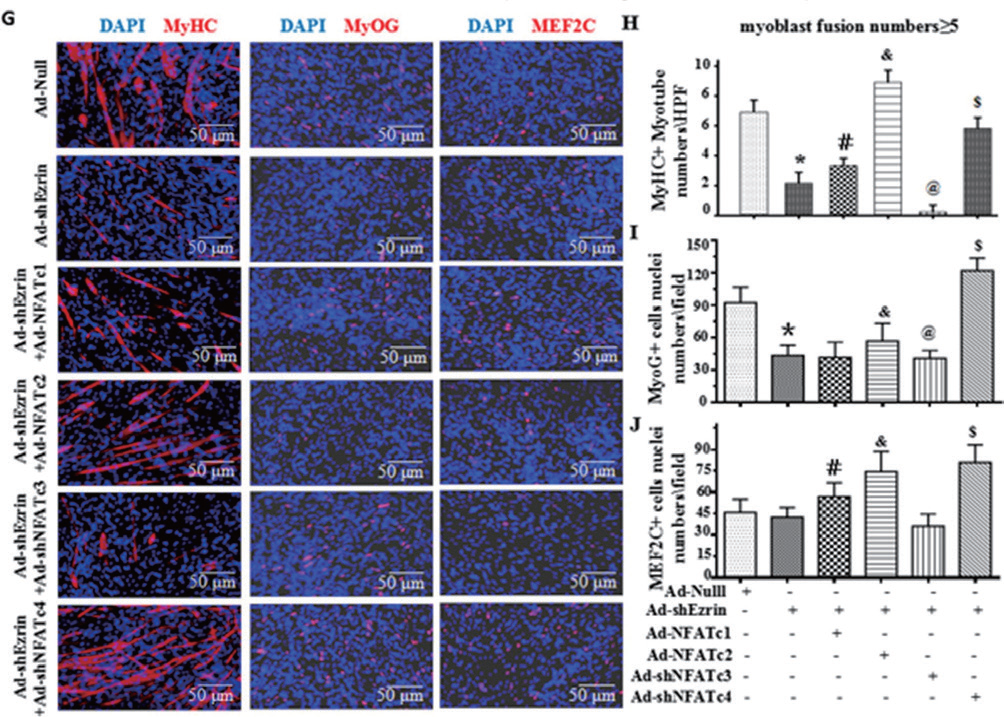
▲ NFAT 信号通路参与Ezrin 介导的成肌细胞分化/ 融合调控 |
1.12 腺病毒感染小鼠脑微血管内皮细胞主要应用:利用腺病毒抑制BMEC 中FGL2 的表达,发现LPS 处理的BMECs 中,TJ 蛋白的降低得到缓解, 表明FGL2 缺乏可能通过上调TJ 蛋白的表达以及恢复母体炎症暴露幼鼠血脑屏障的通透性,来恢复血脑屏 障的功能。此外作者还评估了促炎细胞因子的水平,发现FGL2 缺乏能显著降低幼鼠大脑中促炎细胞因子 的水平。本研究揭示FGL2 可能是治疗产前脑损伤和其他神经炎症疾病的潜在靶点。
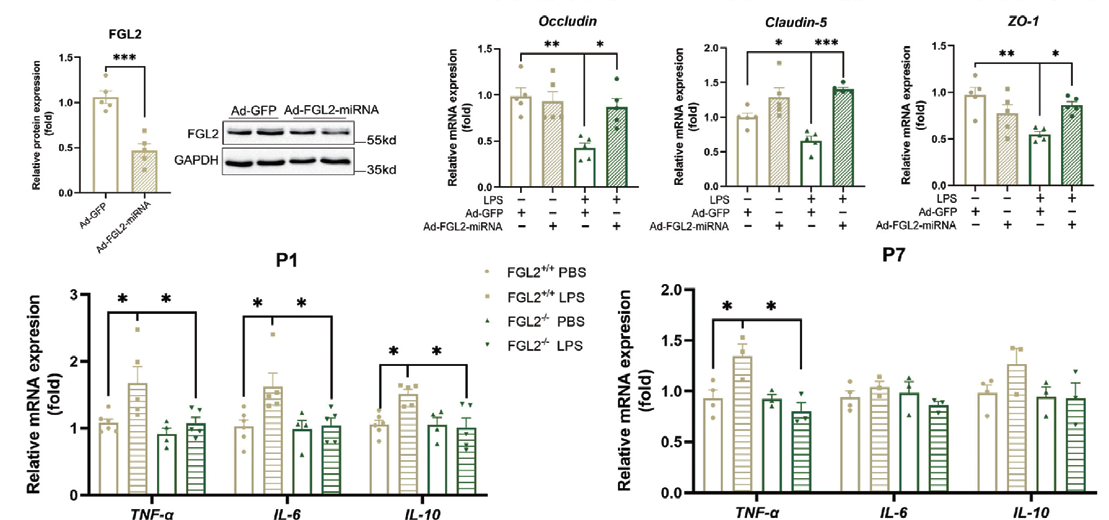
▲ FGL2 缺乏缓解新生小鼠血脑屏障损伤和脑炎症 |
||||||||||||||||||||||
1.13 腺病毒感染小鼠胚胎成纤维细胞主要应用:从纯合子DHX33 lox/lox 小鼠中分离出胚胎成纤维细胞,在MOI 为150 的条件下利用Ad- Cre 处理这些细胞,经过Cre 处理发现DHX33 蛋白以剂量依赖的方式被删除。
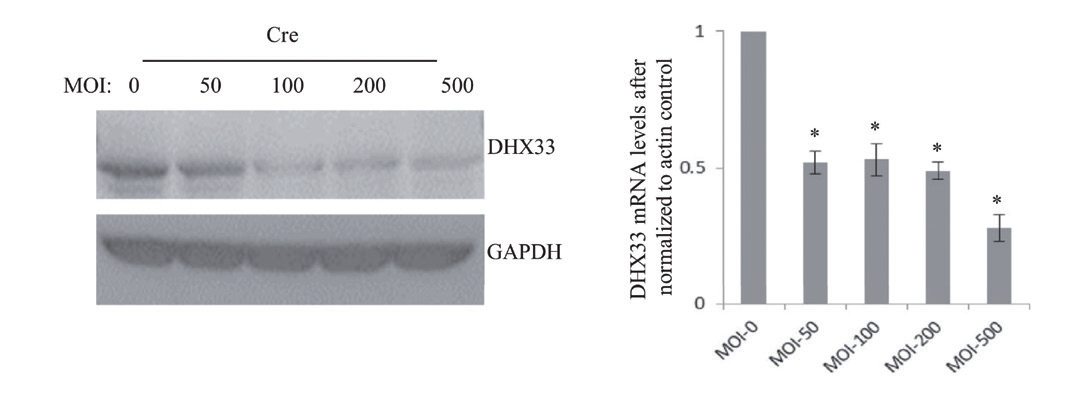
▲ Ad-Cre 转染小鼠胚胎成纤维细胞(MEFs)进行DHX33 的删除 |
|||||||||||||||||||||||
2、腺病毒体内应用策略(部分) |
|||||||||||||||||||||||
2.1 腺病毒 - 立体定向肿瘤内注射 - 大鼠垂体主要应用:将Ad-KBTBD7 经立体定向注射方式注入泌乳素瘤大鼠垂体内,联合CAB(卡麦角林,多巴 胺激动剂)给药,肿瘤不仅明显增重,且DRD2 表达降低以及PRL 水平增加,表明KBTBD7 可以显著增 强雌激素诱导的大鼠垂体原位瘤的DA 抵抗。本研究揭示了DRD2 蛋白降解的新机制,表明KBTBD6/7- DRD2 轴调节PA 对DA 处理的敏感性。因此,KBTBD6/7 可能成为垂体肿瘤的一个有希望的治疗靶点。
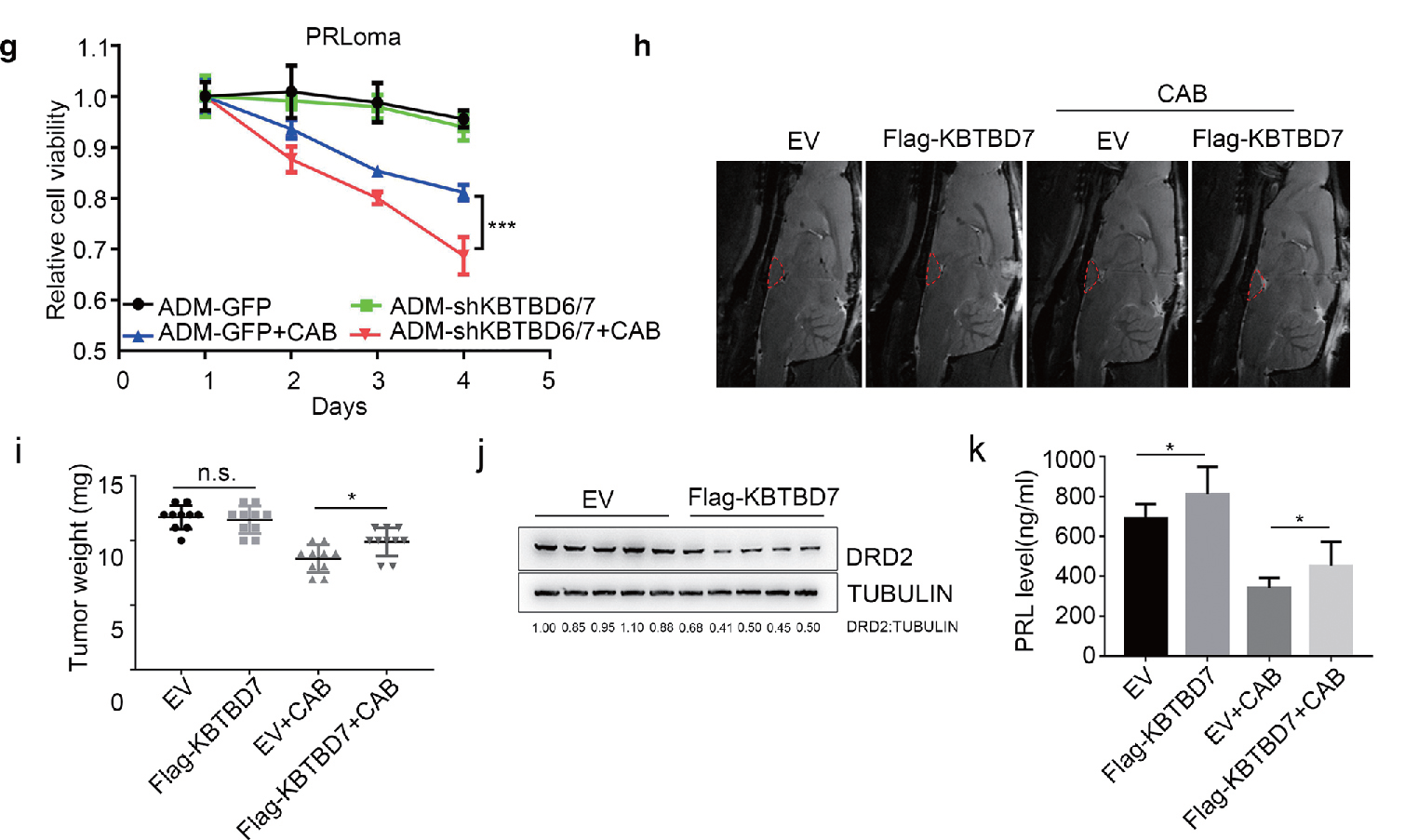
▲ KBTBD7 调节垂体肿瘤对DAs 的敏感性 |
2.2 腺病毒 - 肾皮质内注射 - 小鼠肾脏主要应用:将Ad-UCP1 和Ad-NC 分别注射入小鼠肾脏,构建UCP1 过表达的AKI(急性肾损伤)小鼠模型, UCP1 的过表达明显降低AKI 模型中的脂质含量,显著改善了由顺铂刺激引发的肾形态和肾小管的损伤程 度,肾损伤程度大大降低。本研究首次系统分析了AKI 中脂质组成的变化,发现脂质积累程度与UCP1 的 表达水平呈高度负相关,通过上调UCP1 可以促进AMPK/ULK1/ 自噬通路,减轻AKI 的脂质堆积,进而 显著抑制AKI 发展。该研究为AKI 的治疗提供了新的思路和靶点,为开发新药和联合治疗提供了可能。
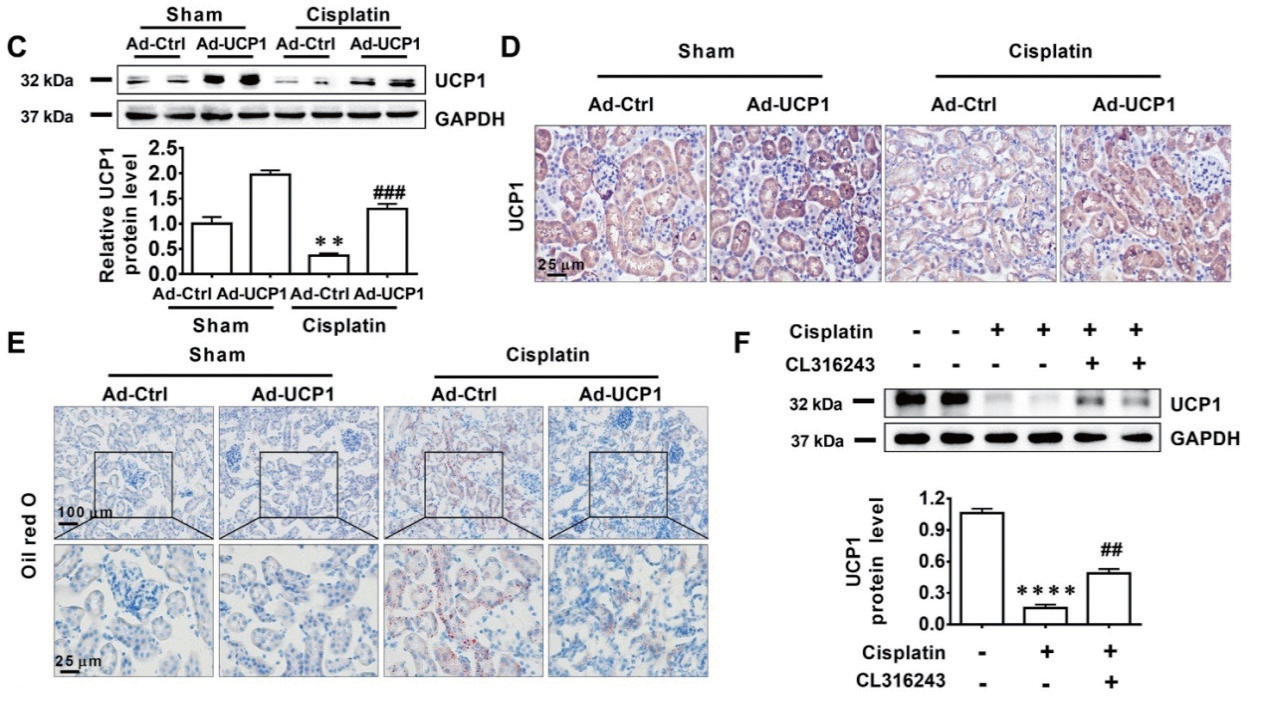
▲ sEVs-pirb 促进GBM 的进展 |
||||||||||||||||||||||
2.3 腺病毒 - 尾静脉注射 - 小鼠肝脏主要应用:肝细胞分泌的FAM3D 调节肝脏糖脂代谢,通过尾静脉注射Ad-FAM3D 在HFD 小鼠中过表 达FAM3D,显著抑制肝脏葡萄糖生成,改善葡萄糖耐受不良和高血糖,减少肝脏脂质沉积。此外,Ad- FAM3D 注射降低了HFD 小鼠血清CHO 水平,但没有降低TG 水平。本研究发现肝细胞分泌的FAM3D 通过激活FPR1-hnRNP U-GR-SCAD 通路增强肝细胞内脂质氧化改善肝脂肪变性,在肥胖情况下,肝脏 FAM3D 表达增加是抵抗糖脂代谢紊乱的一种代偿机制。
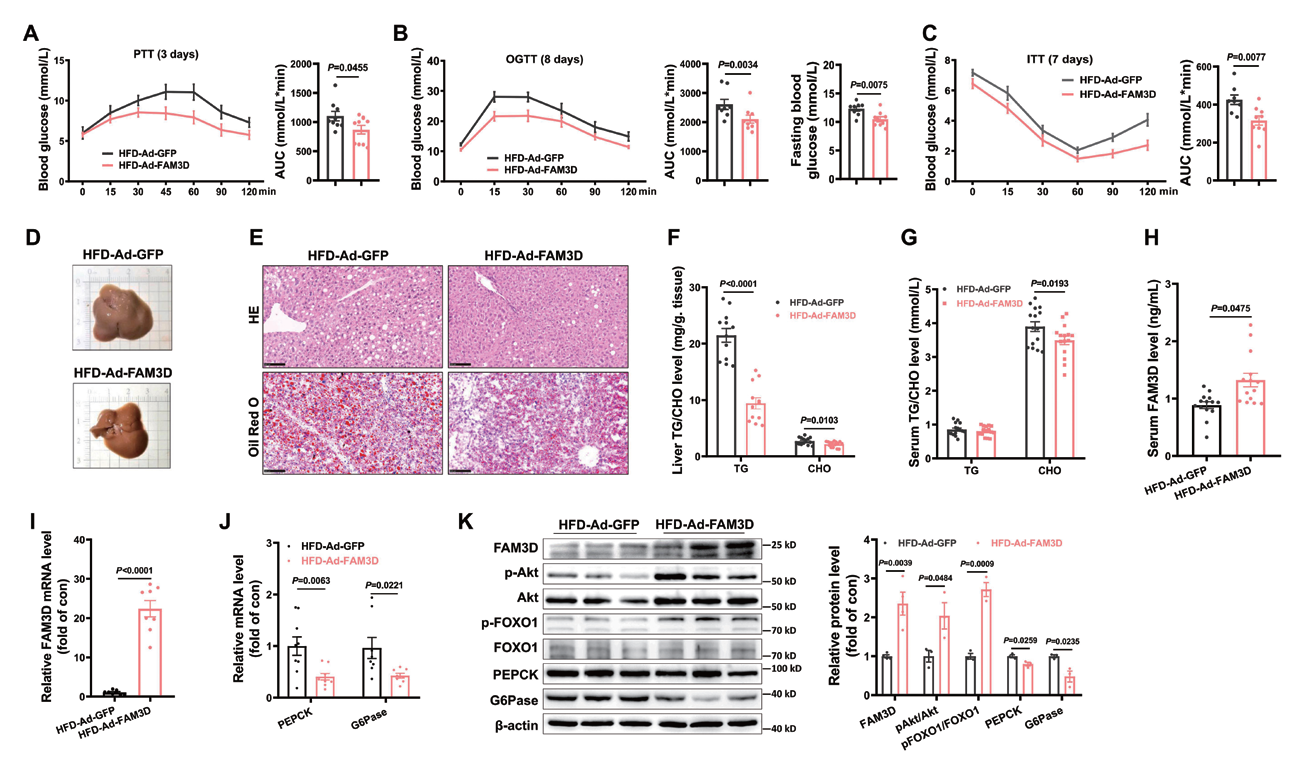
▲ 肝脏FAM3D 过表达可改善HFD 小鼠糖脂代谢失调 |
2.4 腺病毒 - 小鼠子宫主要应用:为确定PES1 是否在子宫内膜异位症的体内发展中发挥作用,作者通过子宫组织自体移植建立 了子宫内膜异位症小鼠模型,并利用Ad-PES1 在小鼠体内过表达Pes1。结果发现过表达Pes1 显著降低 了子宫内膜异位症样病变的重量和体积。此外,与Ad-empty 和PBS 组相比,Ad-PES1 组的TCF21 和 ERβ 蛋白水平降低,表明PES1 在子宫内膜异位症的发展中起着至关重要的作用,过表达PES1 可能是一 种很有前途的治疗方法。
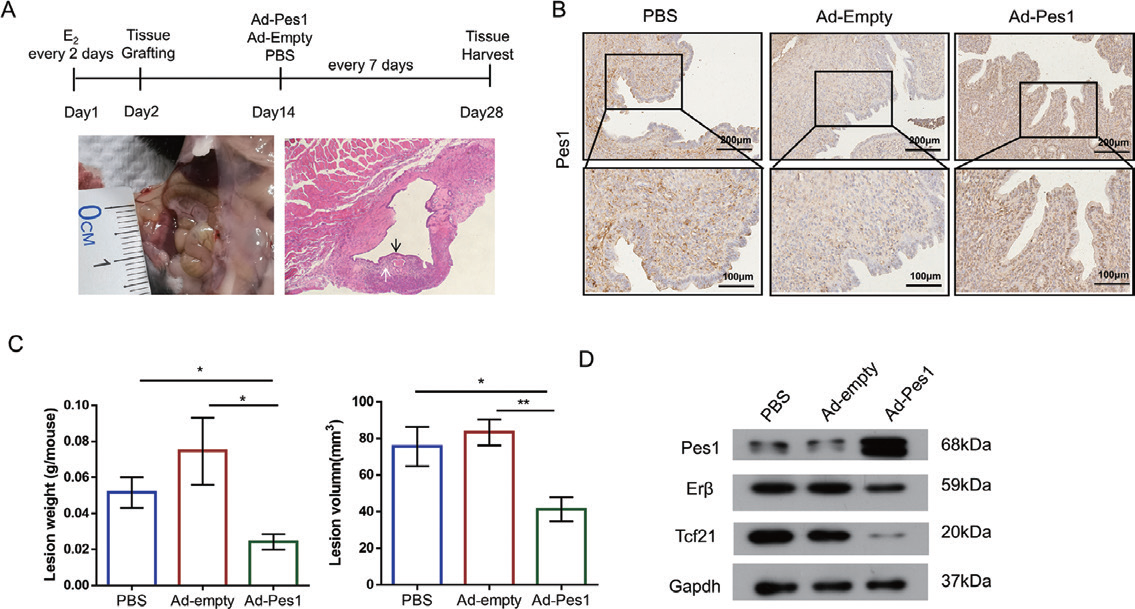
▲ 过表达PES1 对子宫内膜异位症的治疗作用 |
||||||||||||||||||||||
2.5 腺病毒 - 左室壁周围注射 - 大鼠心脏主要应用:利用Ad-JMJD6 实现心脏特异性的JMJD6 过表达,显著减弱ISO 诱导的心肌肥厚特征,同时 JMJD6 过表达还减弱肥厚基因(ANF 和BNP) 和纤维化基因(COL1A1、COL3A1 和CTGF) 的表达,表明 JMJD6 可抑制ISO 诱导的大鼠心脏肥厚和纤维化。本研究发现JMJD6 是一种具有保护心脏功能的去甲基化 酶,通过在p65 的R149 位点去甲基化减少NF-κB 信号传导,从而改善ISO 诱导的病理性心肌肥大,揭 示靶向JMJD6 可能是病理性心肌肥厚的潜在治疗靶点。
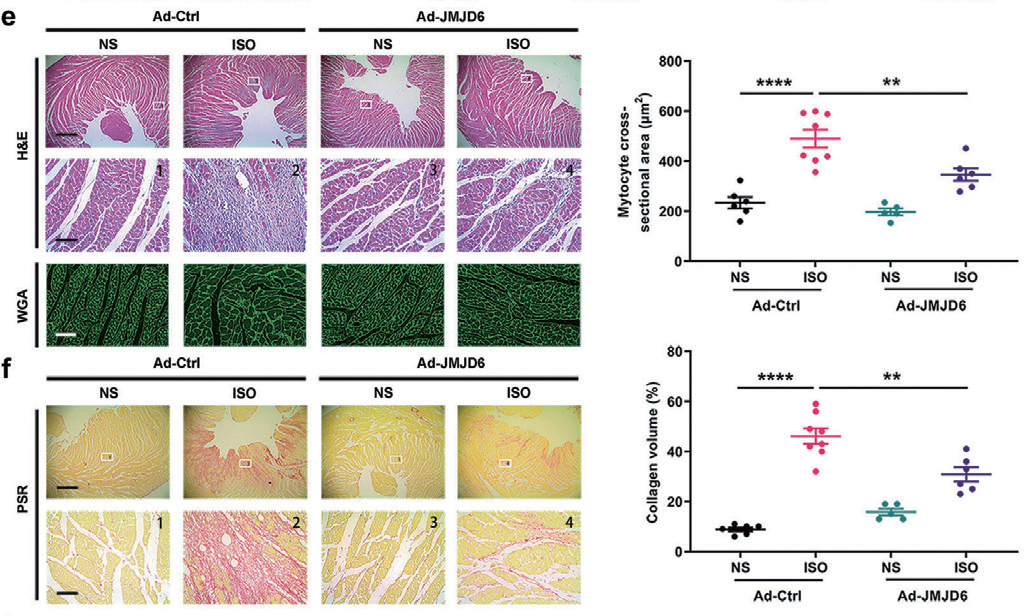
▲ KBTBD7 调节垂体肿瘤对DAs 的敏感性 |
2.6 腺病毒 - 颈总动脉注射 - 大鼠心血管主要应用:利用腺病毒干扰载体下调颈动脉中的Beclin1,明显改善了球囊损伤后TMAO(三甲胺N- 氧化物) 诱导的自噬损害,并逆转了再内皮化率的降低。本研究首次揭示球囊损伤后TMAO 通过Beclin1 介导的 自噬流量损伤加重血管新生内膜增生,因此调节TMAO 代谢可能是未来各种心血管疾病中对抗血管重塑 的一种有前景的策略。
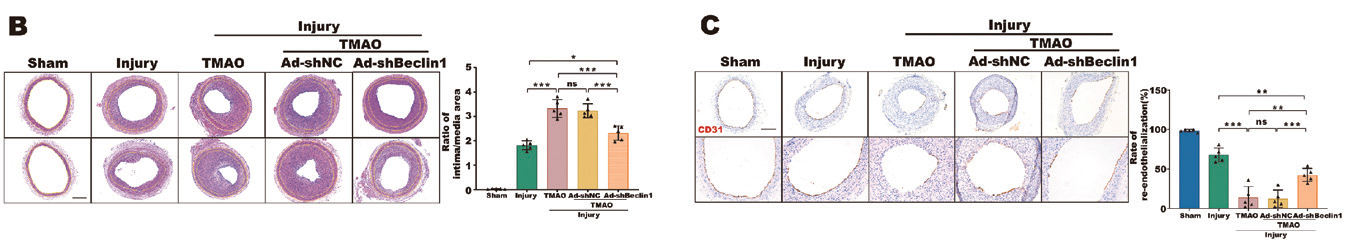
▲ Beclin1 的下调恢复了自噬通量,减弱了TMAO 诱导的内膜增生 |
||||||||||||||||||||||
2.7 腺病毒 - 瘤内注射 - 小鼠结肠主要应用:通过注射表达人CSF1 的重组腺病毒载体,建立了CSF1 过表达的HT-29 异种移植瘤小鼠模型, CSF1 的过表达明显增加了肿瘤浸润的巨噬细胞数量,提高小鼠肿瘤组织中IL-8 mRNA 水平,并且显著减小肿瘤体积。本研究揭示CSF1 是结肠癌微环境中的一个重要因子,参与了结肠癌细胞与肿瘤浸润的巨噬 细胞之间的相互作用。
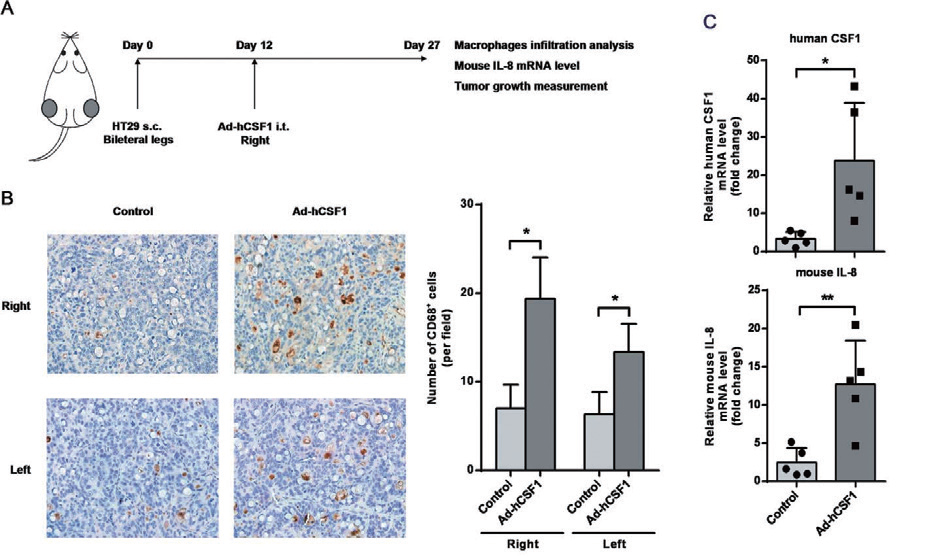
▲ 瘤内CSF1 过表达影响体内巨噬细胞浸润和结肠癌肿瘤生长 |
2.8 腺病毒 - 鼻吸注射 - 小鼠肺部主要应用:将DHX33 lox/lox 小鼠与K-Ras (G12D-LSL) 小鼠杂交,产生了四种不同基因型的后代。将 Ad-Cre 以鼻吸的方式给药至四种小鼠,结果表明在Ras 驱动的肺癌模型中,DHX33 基因缺失可阻断肺肿瘤的发生。本研究首次证实了DHX33 在体内Ras 驱动型肺癌发展中的关键作用,并强调了以DHX33 为靶点的药物治疗Ras 突变型肺癌是一种可行的选择。
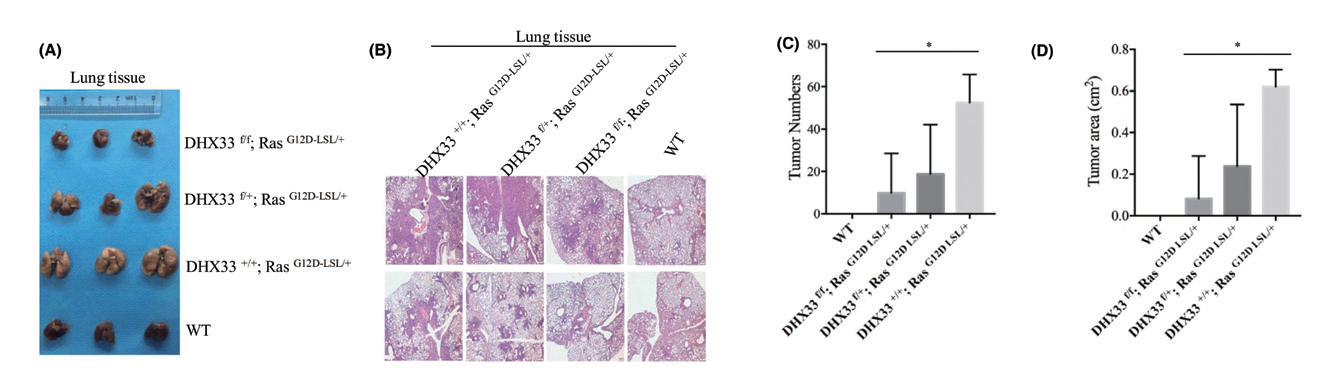
▲ Beclin1 的下调恢复了自噬通量,减弱了TMAO 诱导的内膜增生 |
||||||||||||||||||||||
维真生物采用的包装系统包括了AdEasy和AdMax系统。其中,AdMax系统操作简便、重组效率高、获得的病毒产率也高,故成为目前腺病毒包装主要采用的系统。下图是AdEasy和AdMax系统的示意图:
|
|
| AdEasy系统 |
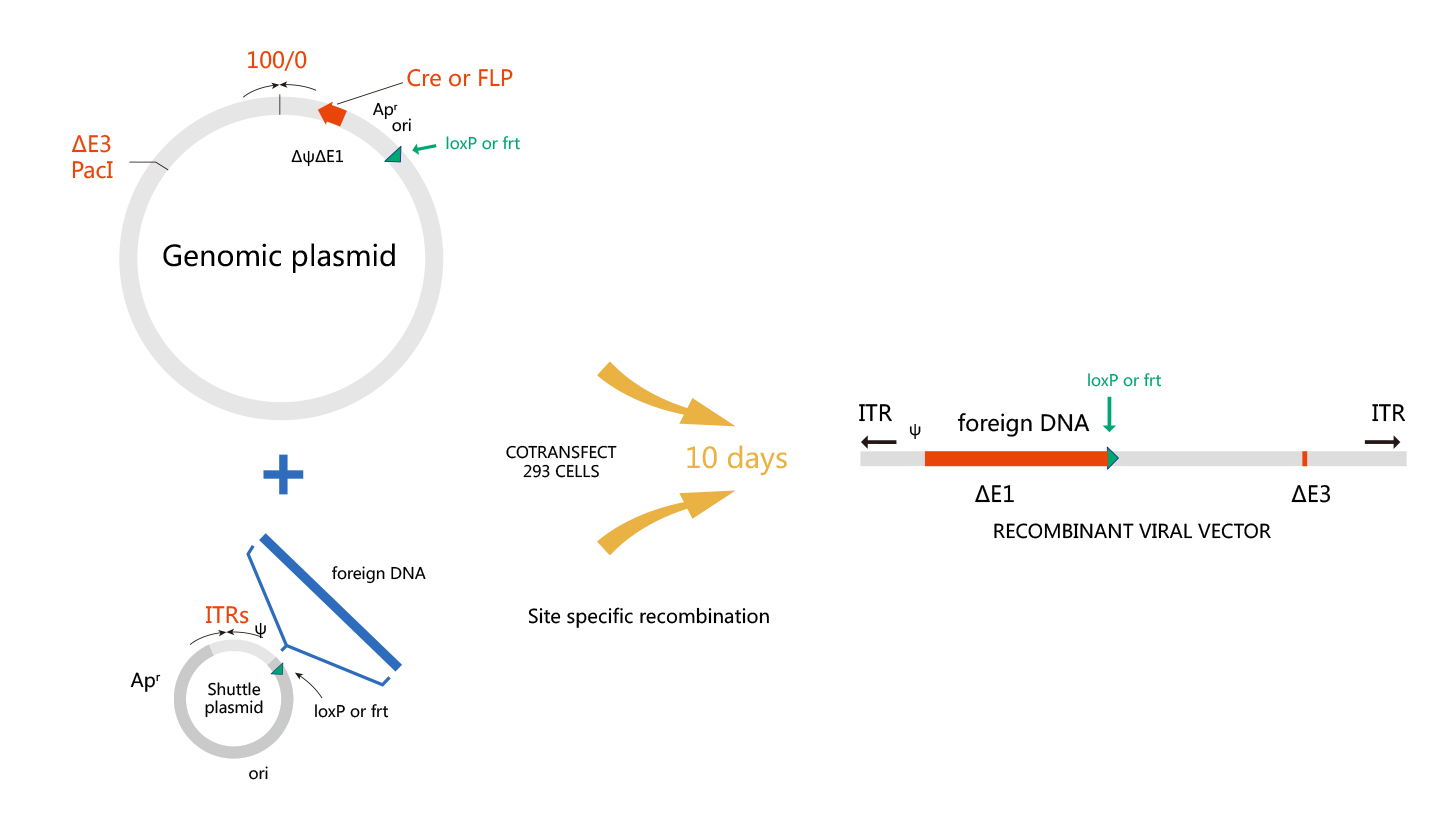 |
| AdMax系统 |
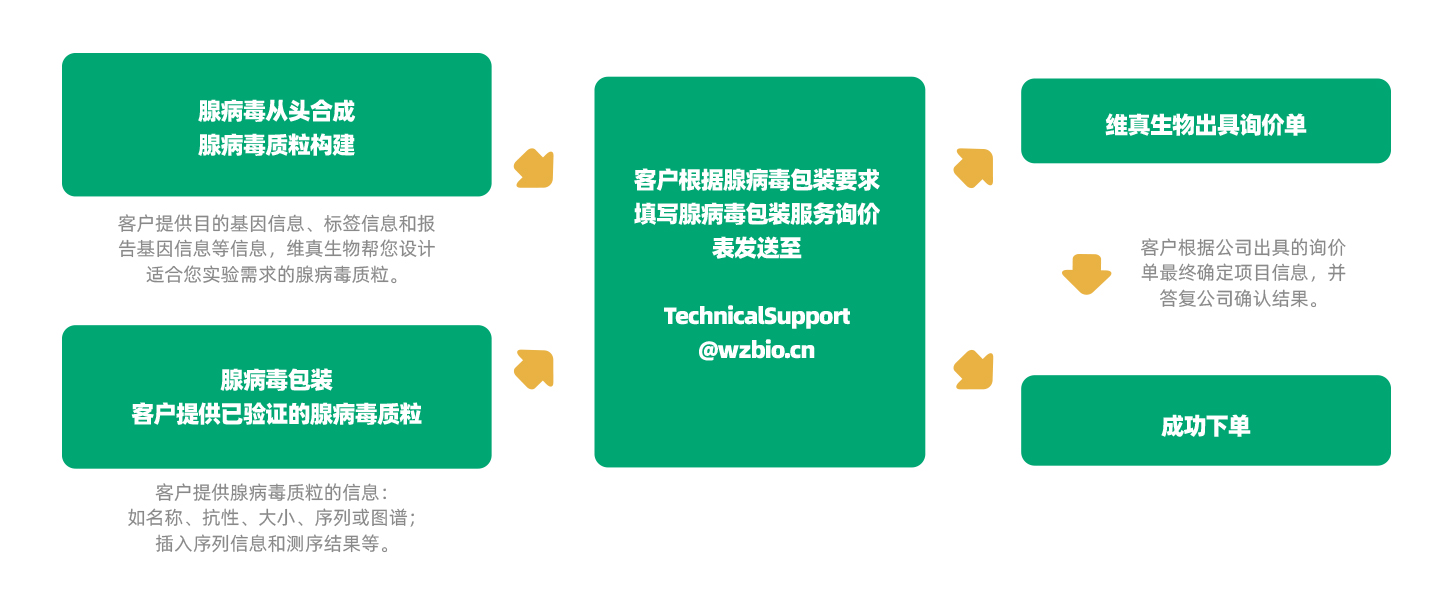
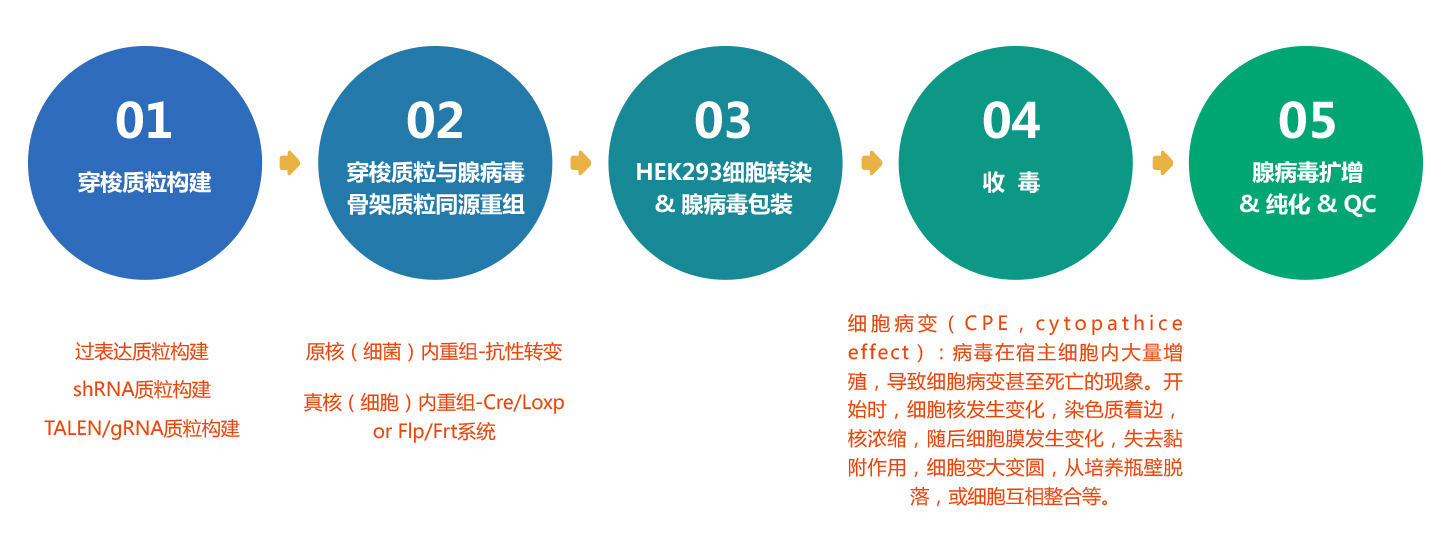
维真生物可以提供从腺病毒(AdV)质粒设计、构建、包装到表达分析的所有服务外包项目。而且,维真生物全套的腺病毒载体和表达系统可用于体内和体外表达human/mouse/rat ORFs,lncRNA,circRNA,shRNAs和CRISPR/gRNA。维真生物生产的腺病毒具有滴度高、纯度高和稳定性高的三高优点。下面是腺病毒从头合成的流程:
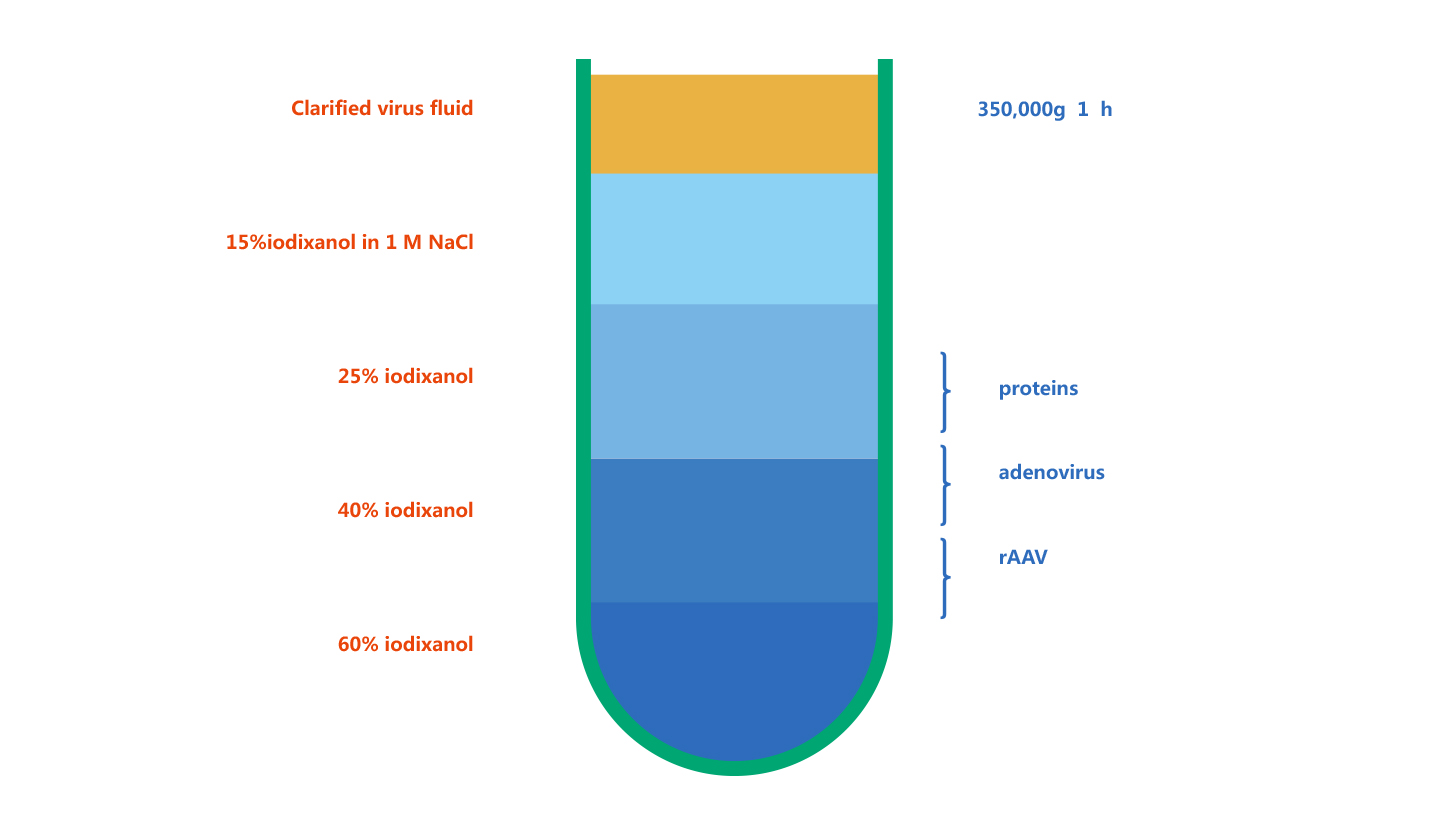
维真生物采用碘克沙醇密度梯度超速离心法对AdV病毒进行纯化。碘克沙醇是一种造影剂,已接受了临床检验。它具有非离子型、对细胞无毒和代谢惰性等优点。现在在病毒纯化方面应用广泛。它利用不同颗粒之间存在的沉降系数差,在一定离心力作用下,颗粒各自以一定速度沉降,在密度梯度不同区域上形成区带。下图是AdV病毒纯化后的示意图:
维真生物的AdV病毒采用多种方法来定量,主要包括QPCR法、孔稀释法和快速腺病毒滴度免疫检测法。下表是各方法的详情:
|
滴度标定法 |
原理 |
滴度单位 |
用途 |
|
QPCR法 |
用腺病毒基因组特异性引物进行QPCR。在定量曲线的线性范围内,Ct值与已知拷贝数质粒Ct值的比值,即为病毒基因组的初始拷贝数 |
vp/ml |
预制腺病毒库中 |
|
孔稀释法 |
数荧光 |
pfu/ml |
带荧光腺病毒的 |
|
免疫法 |
通过抗腺病毒抗体与感染了腺病毒供试样品的细胞结合,再用辣根过氧化物酶标记的二抗与一抗结合,经显色液显色,被感染的细胞显示出明显的棕色,通过计数及计算,测定腺病毒滴度 |
pfu/ml |
该方法不受荧光限制,适用于所有腺病毒滴度的测定 |
|
↑↑ 点击返回,查看其它部分 ↑↑ |
|
1. 载体全: |
多种表达载体、多种标签载体和多种报告基因可供客户选择。 |
|
2. 货期短: |
Human ORF cDNA克隆可在2-3天穿梭至其他标签(3×Flag,Myc,Flag,HA,6×His,Flag & His,GFP,RFP等)的穿梭载体,个性化定做腺病毒; |
|
3. 纯度高: |
采用改良的碘克砂醇密度梯度超速离心法进行腺病毒纯化,对于动物没有任何毒性; |
|
4. 服务优: |
可根据客户具体实验,由公司专业技术人员提供载体构建方案,提供特殊定制服务。 |
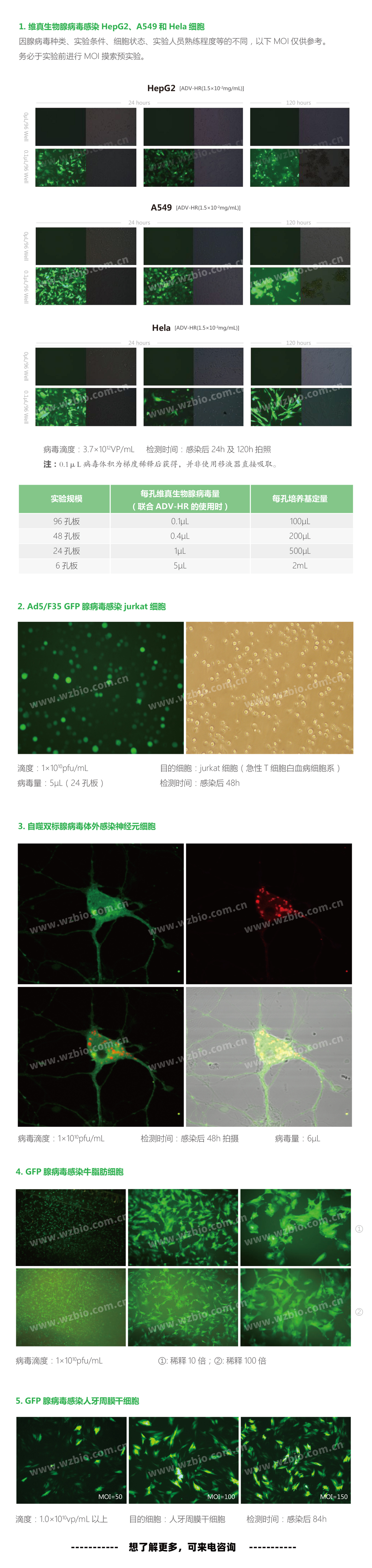 |
1. 您好,我感染的细胞是滋养层细胞,感染之前在六孔板中接种十万细胞,贴壁16h后加加入病毒(MOI=40),30h后细胞长满。我想咨询:病毒感染后实验组跟对照组相比细胞生产差不多,正常吗?
如果感染腺病毒的细胞状态与未感染组的细胞状态无明显差异,说明该病毒对细胞没有明显毒性作用。因此,未必是病毒数量不够。非腺病毒包装细胞被腺病毒感染后是不会有什么明显的特征。如果您在加入病毒的时候是提前把病毒用培养基稀释混匀之后再加入培养皿中的,那么高MOI值(导致细胞飘起来)对于目的细胞是比较高的。因此,您不用担心,可以先检测一下是否有蛋白表达。
2. 腺病毒感染细胞后是否需要更换新鲜培养液?
是否需要更换新鲜培养液需要看加入病毒的量,病毒量多的时候会对细胞有毒性:如果镜检时看到细胞状态有变化,需要在4-8h左右更换新鲜培养液;如果镜检时看到细胞状态无明显变化,那么病毒感染细胞之后就无需更换新鲜培养液。或者,如果您担心病毒长时间作用于细胞会有毒性,也可在12h后更换新鲜培养液。
3. 腺病毒感染细胞4-6h之后发现细胞全部飘起来了,请问这是怎么回事?
造成上述现象的原因主要有三点:
1)细胞的状态:感染之前铺板的细胞一定要使用健康的细胞;
2)实验操作不当:在加入病毒液的时候,如果局部病毒浓度过高会导致细胞死亡。因此,我们建议将病毒和培养基混匀之后再加入细胞中。
3)如果以上2点均无误,那就是加入的病毒量太大了。
4. 腺病毒感染细胞时,加入病毒量的依据是什么?
要得到最佳实验结果,确定腺病毒的最适用量是关键。量不足达不到100%感染效率;量太高则会对细胞产生毒性。那么如何确定呢?"感染复数"(MOI,multiplicity of infection)起着决定作用。MOI是是感染时病毒与细胞数量的比值。不同细胞系细胞表面的腺病毒受体数量不同,这就决定了不同细胞系的MOI会有所不同。通常,易感染细胞系所使用的MOI范围为10-100。 但是,对于某些难感染的细胞系,MOI可能需要高达1000。对于大多数细胞系来说,最佳MOI的范围很窄。我们建议您查阅文献或者在正式实验前,在目的细胞中用报告基因的腺病毒进行预实验摸最佳MOI。
最适病毒用量的计算公式:
病毒用量pfu=最佳MOI×细胞数目/病毒滴度
例如, 如果您目的细胞的最佳MOI=10,您需要感染106的细胞,那么您共计需要107pfu的病毒. 如果病毒滴度为1×1010 pfu/mL, 那么您实验需要的病毒量就是1ul.
5. 腺病毒感染细胞之后多久可以从基因水平和蛋白水平检测表达?如果蛋白是分泌性蛋白和非分泌性蛋白,检测时间是否有区别?
腺病毒携带的目的基因表达快。如果您想从基因水平检测基因的表达,那么在病毒感染细胞12-24h之间进行检测;如果您想从蛋白水平检测基因的表达,那么在病毒感染细胞48-72h之间进行检测。
对于分泌性蛋白和非分泌性蛋白,检测时间一样。如果担心蛋白分泌出来,也可以将培养基也一起带着做western blot。
6. 腺病毒毒种被细菌污染了,没有多余毒种,也没有构建好的载体,怎么办?
因为腺病毒的直径大小是90-100nm,而细菌的直径大小要大的多,可以用22um的滤器进行过虑。如果您的病毒量很低,担心过虑后损失太多,无法进行后面的扩增工作,您可以先用污染了细菌的腺病毒直接扩增,当然扩增的结果是得到了污染了细菌的腺病毒,但病毒的量会提高一些,这样再用22um的滤器进行过滤。
7. 如何鉴定腺病毒递送的目的基因?
可以按照如下方法进行鉴定:
①针对基因设计引物,同时作为扩增和测序引物;
②将病毒用蛋白酶K处理后做为模板,如果是未纯化的病毒还需要过纯化柱处理掉培养基的成分;
③以处理过的病毒为模板进行PCR;
④用PCR引物对PCR产物测序。
8. 慢病毒、腺病毒、腺相关病毒三种病毒有什么区别?如何选择?
对于转染困难的细胞,科研工作者通常会选择病毒来导入目的基因。目前,腺病毒、慢病毒和腺相关病毒是常用的病毒载体工具。那么如何选择适合您实验体系的病毒工具,请参考下面3种病毒工具的比较:
|
病毒表达系统 |
腺病毒 |
腺相关病毒 |
慢病毒 |
|
病毒基因组 |
dsDNA |
ssDNA |
ssRNA |
|
病毒外壳 |
无 |
无 |
具有包膜蛋白 |
|
基因组大小 |
38-39kb |
5kb |
9kb |
|
包装容量 |
7.5kb |
4.7kb |
6kb |
|
感染的细胞类型 |
分裂细胞和非分裂细胞 |
分裂细胞和非分裂细胞 |
分裂细胞和非分裂细胞 |
|
整合至宿主基因组 |
非整合 |
非整合 |
整合 |
|
表达丰度 |
高水平表达 |
高水平表达 |
中到高水平表达 |
|
表达时间 |
快(1-2天) |
1-2周(体内) |
慢(2-4天) |
|
外源基因持续表达时间 |
短暂 |
潜在的持久 |
长久 |
|
免疫反应 |
较高 |
极低 |
低 |
|
相对病毒滴度 |
10E10pfu/mL |
10E13VG/mL |
10E8TU/mL |
|
生物安全等级 |
BSL-2 |
BSL-2 |
BSL-2 |
|
↑↑ 点击返回,查看其它部分 ↑↑ |
ORF现货腺病毒:
|
1. |
||
|
2. |
||
|
3. |
||
|
4. |
||
|
5. |
||
|
6. |
||
|
7. |
||
|
8. |
||
|
9. |
||
|
10. |
||
|
11. |
||
|
12. |
||
|
13. |
MicroRNA现货腺病毒:
|
1. |
||
|
2. |
基因过表达方面:
|
1. |
||
|
2. |
||
|
3. |
J Cell Physiol. (IF=5.546) . Wang M, et al. (2021) . Lack of Mof reduces acute liver injury by enhancingtranscriptional activation of Igf1. [过表达/干扰 急性肝损伤] |
|
|
4. |
Front Cell Dev Biol. (IF=5.201) . Gu J, et al. (2021) . CFTR Deficiency Affects Glucose Homeostasis via Regulating GLUT4 Plasma Membrane Transportation. [过表达 成肌细胞] |
|
|
5. |
Front Immunol. (IF=5.085) . Wu L, et al. (2021) . EZH2 Inhibition Interferes With the Activation of Type I Interferon Signaling Pathway and Ameliorates Lupus Nephritis in NZB/NZW F1 Mice. [过表达 狼疮肾炎] |
|
|
6. |
[4]Oxid Med Cell Longev. (IF=5.076) . Gao Y, et al. (2021) .Rap1GAP Mediates Angiotensin II-Induced Cardiomyocyte Hypertrophy by Inhibiting Autophagy and Increasing Oxidative Stress. [过表达 心肌] |
|
|
7. |
J Cell Mol Med. (IF=4.486) . Lei W,et al. (2021) . MARCH5 restores endothelial cell function against ischaemic/hypoxia injury via Akt/eNOS pathway. [过表达 心肌] |
|
|
8. |
J Biol Chem. (IF=4.238) . Zhang Z,et al. (2021) . The adaptor protein GIPC1 stabilizes the scavenger receptor SR-B1 and increases its cholesterol uptake. [过表达/干扰 肝脏] |
|
|
9. |
J Anim Sci Biotechnol. (IF=4.167) . Liu J, et al. (2021) . Comprehensive evaluation of the metabolic effects of porcine CRTC3 overexpression on subcutaneous adipocytes with metabolomic and transcriptomic analyses. [过表达 脂肪细胞] |
|
|
10. |
Aging. (IF=4.831) .Chen Y, et al. (2020). Mitophagy impairment is involved in sevoflurane-induced cognitive dysfunction in aged rats[自噬双标 术后认知功能障碍]. |
|
|
11. |
||
|
12. |
||
|
13. |
||
|
14. |
||
|
15. |
||
|
16. |
||
|
17. |
||
|
18. |
||
|
19. |
||
|
20. |
||
|
21. |
||
|
22. |
||
|
23. |
||
|
24. |
基因沉默方面:
|
1. |
||
|
2. |
||
|
3. |
||
|
4. |
||
|
5. |
||
|
6. |
||
|
7. |
基因编辑方面:
|
1. |
||
|
2. |
||
|
3. |
|
↑↑ 点击返回,查看其它部分 ↑↑ |

400-077-2566

service@wzbio.cn